Summary of Berberine
Primary Information, Benefits, Effects, and Important Facts
Berberine is an alkaloid extracted from various plants used in traditional Chinese medicine.
Berberine is supplemented for its anti-inflammatory and anti-diabetic effects. It can also improve intestinal health and lower cholesterol. Berberine is able to reduce glucose production in the liver. Human and animal research demonstrates that 1500mg of berberine, taken in three doses of 500mg each, is equally effective as taking 1500mg of metformin or 4mg glibenclamide, two pharmaceuticals for treating type II diabetes. Effectiveness was measured by how well the drugs reduced biomarkers of type II diabetes.
Berberine may also synergize with anti-depressant medication and help with body fat loss. Both of these benefits need additional evidence behind them before berberine can be recommended specifically for these reasons.
Berberine’s main mechanism is partly responsible for its anti-diabetic and anti-inflammatory effects. Berberine is able to activate an enzyme called Adenosine Monophosphate-Activated Protein Kinase (AMPK) while inhibiting Protein-Tyrosine Phosphatase 1B (PTP1B).
Berberine has a high potential to interact with a medications, and some interactions may be serious.
Berberine is one of the few supplements in the Examine.com database with human evidence that establishes it to be as effective as pharmaceuticals.
Things to Know
Do Not Confuse With
Piperine (Black Pepper extract), Berberol (Brand name), Berberrubine (Metabolite)
Things to Note
- High doses of berberine taken acutely, due to their poor intestinal uptake rate, may cause cramping and diarrhea; for this reason, berberine should be taken in multiple doses throughout the day
- Berberine is known to inhibit CYP2D6, CYP2C9, and CYP3A4, which can lead to a host of drug interactions, some of which can be serious
- Berberine is known to induce the protein concent of P-glycoprotein
- Berberine interacts with organic anion transporter proteins, which may limit tissue uptake of metformin
- Berberine may interact with microlide antibiotics such as azithromycin and clarithromycin at hERG channels on the heart, leading to serious cardiotoxicity
Is used for
Also used for
Is a form of
Goes Well With
- P-glycoprotein (P-Gp) inhibitors increase absorption rate, with Milk Thistle demonstrated in humans and Stephania tetrandra being promising
- Sodium caprate (increases absorption, not related to P-Glycoprotein)
- Atrogin-1 inhibition (theoreticallyreverses the possible degradation of lean mass associated with AMPK activation into synthesis)
Does Not Go Well With
- Phosphodiesterase inhibitors (can attenuate but not abolish the increase in cAMP that PDE inhibitors result in, and may reduce their fat-burning effects)
Caution Notice
Known to interact with enzymes of Drug Metabolism. Also may interact with microlide antibiotics such as azithromycin and clarithromycin at hERG channels on the heart, leading to serious cardiotoxicity.
How to Take
Recommended dosage, active amounts, other details
The standard dose of berberine is 900-2,000mg a day, divided into three to four doses.
Berberine should be taken with a meal, or shortly after, to take advantage of the blood glucose and lipid spike associated with eating.
Too much berberine at once can result in stomach upset, cramping, and diarrhea.
Editors' Thoughts on Berberine
Yeah, so I guess I'll just use this area to talk about the 'bad' associated with this supplement. Consider it a panacea or at least inert except for the following (hey, nothing is perfect):
Kurtis Frank
Frequently Asked Questions
Questions and answers regarding Berberine
Q: 5 supplements (and foods) for a stronger heart
A: Cardiovascular diseases (CVDs) are the #1 cause of death globally. But a mix of the right foods and complementary supplements can help decrease your risk factors.
Read full answer to "5 supplements (and foods) for a stronger heart"
A: Cardiovascular diseases (CVDs) are the #1 cause of death globally. But a mix of the right foods and complementary supplements can help decrease your risk factors.
Read full answer to "5 supplements (and foods) for a stronger heart"
Human Effect Matrix
The Human Effect Matrix looks at human studies (it excludes animal and in vitro studies) to tell you what effects berberine has on your body, and how strong these effects are.
Scientific Research
Table of Contents:
- 1 Sources and Structure
- 1.1 Sources
- 1.2 Structure
- 1.3 Berberine Complexes
- 1.4 Related Structures
- 2 Pharmacology
- 2.1 Absorption
- 2.2 Distribution
- 2.3 Metabolism
- 2.4 Excretion
- 2.5 Enzymatic Interactions
- 3 Molecular Targets
- 3.1 AMPK
- 4 Longevity and Life Extension
- 4.1 Telomeres
- 5 Neurology
- 5.1 Pharmacokinetics
- 5.2 Adrenergic Neurotransmission
- 5.3 Serotoninergic Neurotransmission
- 5.4 Dopaminergic Neurotransmission
- 5.5 Memory and Learning
- 5.6 Depression
- 5.7 Sedation
- 5.8 Analgesia
- 5.9 Alzheimer's Disease
- 5.10 Neuroprotection
- 5.11 Addiction and Dependence
- 6 Cardiovascular Health
- 6.1 Cardiac Tissue
- 6.2 Triglycerides
- 6.3 Blood Flow
- 6.4 Platelets
- 6.5 Atherosclerosis
- 7 Fat Mass and Obesity
- 8 Interactions with Glucose Metabolism
- 8.1 Absorption
- 8.2 Interventions
- 9 Skeletal Muscle and Physical Performance
- 10 Skeleton and Bone Metabolism
- 10.1 Calcitriol
- 10.2 Joint Inflammation
- 11 Inflammation and Immunology
- 11.1 Inflammation (General)
- 11.2 Macrophages
- 11.3 T-Cells
- 11.4 Virology
- 12 Interactions with Oxidation
- 12.1 Endoplasmic Reticulum
- 12.2 Lipid Peroxidation
- 13 Interactions with Hormones
- 13.1 Testosterone
- 13.2 Estrogen
- 13.3 Leptin
- 13.4 GLP-1
- 14 Cancer Metabolism
- 14.1 Mechanisms (General)
- 14.2 Proliferation and Angiogenesis
- 14.3 Migration and Metastasis
- 14.4 Autophagy
- 14.5 Adjunct Therapy Potential
- 14.6 Brain
- 14.7 Breast
- 14.8 Liver
- 14.9 Oral
- 14.10 Thyroid
- 14.11 Colon
- 14.12 Prostate
- 14.13 Immunological Cancers
- 15 Interactions with the Liver (Hepatology)
- 15.1 Glucose Interactions
- 15.2 Lipids and Cholesterol
- 15.3 Fibrosis
- 15.4 Liver Enzymes
- 15.5 Nonalcoholic Fatty Liver Disease
- 16 Interactions with the Intestinal Tract
- 16.1 Interactions with Glucose Metabolism
- 16.2 Diarrhea
- 16.3 Ulcerative Colitis
- 16.4 Colonic Microflora
- 17 Interactions with other Organ Systems
- 18 Other Possible Therapeutic Roles
- 19 Nutrient-Nutrient Interactions
- 19.1 Metformin
- 19.2 Statins
- 19.3 Policosanol and Red Yeast Extract
- 19.4 Sodium Caprate
- 19.5 Berberol
- 19.6 Er-Xian
- 20 Safety and Toxicology
- 20.1 General
- 20.2 Contraindictions
1Sources and Structure
1.1. Sources
Berberine (2,3-methylenedioxy-9,10-dimethoxy-protoberberine) has been used historically in Ayurvedaand Traditional Chinese medicine (vicariously through herbs which contain it) as an anti-microbial, anti-protozoal, and anti-diarrheal agent.[4]
It has shown efficacy against various bacteria strains such as cholera, giardia, shigella, and salmonella; potentially also staphylococcus, streptococcus, and clostridium.[4][5] Its actions against protozoa extend to Giardia lamblia, Trichomonas vaginalis, Leishmania donovani, and Malaria.[6][7][8]Surprisingly, crude extracts are more potent than isolated berberine in these anti-protozoan effects suggesting synergistic or additive effects with other compounds in these plants.[9]
Various effects against bacteria and protozoa, which underlies a fair bit of traditional usage of the plants containing Berberine
Berberine has been isolated from various plant families including Papaveraceae, Berberidaceae, Fumariaceae, Menispermaceae, Ranunculaceae, Rutaceae, and Annonaceae[10]
- (Berberidaceae family) Berberis Aristata (Tree Turmeric or Indian Barberry)[4] at 5% of the roots or 4.2% of stem and bark.[11] The Berberis genera that also includes Berberine include vulgaris (1.24%), petiolaris (0.43%) as well as thunbergii, aquifolium, and asiatica.[4] Other plant sources of Berberine in this family include Caulis mahoniae[11] and Mahonia aquifolium (Oregon Grape)
- (Rutaceae family) Phellodendron Amurense (Amur Cork Tree, Huang Bai)[4]
Appears to be a common alkaloid present in a variety of plant families, and most of these plants have traditionally been used for digestion or glucose/diabetes related issues in traditional medicine
1.2. Structure
Berberine is known as an isoquinoline alkaloid with the prolonged name of 2,3-methylenedioxy-9,10-dimethoxy-protoberberine.[14] It has a molecular weight of 336.36122g/mol, and is bright yellow in color when isolated.[15]
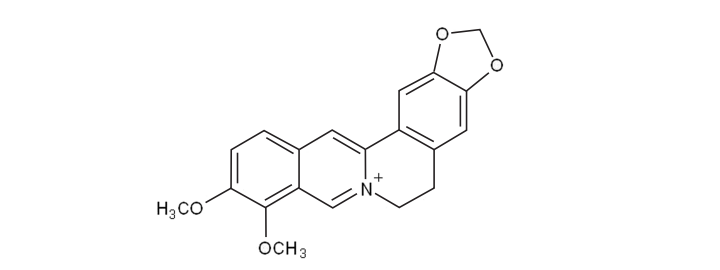
1.3. Berberine Complexes
Berberine can complex at a 1:1 ratio with the flavonoid baicalin (and to a degree, wogonoside),[16] the complex of which can be formed when Radix Scutellariae and Rhizoma Coptidis (sources of Baicalin and Berberine, respectively) are mixed which occurs in a few Traditional Chinese Medicinecombination therapies. It is thought that these complexes (and another Berberine–glycyrrhizin complex) enhance absorption through forming ion-pairs and enhancing fat solubility, where the glucuronide of Baicalin has its carboxylate ion bind to the quaternary ammonium ion of Berberine.[16]
Berberine, due to its quaternary ammonium ion, can form complexes with other compounds also present in Chinese herbal decoctions; these may have different properties or absorption than Berberine per se
1.4. Related Structures
A related protoberberine compound, Dihydroberberine, appears to have similar effects to Berberine but with lower doses (thus, higher potency), with one study suggesting 560mg/kg Berberine had similar effects to 100mg/kg Dihydroberberine in a high-fat fed rat model measuring adiposity and glucose tolerance.[17] This study noted that dihydroberberine was detected in plasma (with a calculated bioavailability of 2.85%) at a dose that Berberine was not,[17] but noted that Berberine may not have practically relevant improved absorption since it readily converts to Berberine in acidic environments (such as the stomach).[18] The researchers then synthesized 8,8-dimethyldihydroberberine as a pharmaceutical alternative.[18]
Dihydroberberine is also found naturally occurring in some plants (such as Arcangelisia flava[19] and Coptidis Chinensis[20])
Dihydroberberine, although less studied than Berberine itself, appears to be more effective on the main parameter of Berberine inducing AMPK when used in vitro; may merely convert to Berberine in practical situations
The synthetic deriviative known as CPU86017 (p-chlorobenzyltetrahydroberberine chloride or Raisanberine) appears to be a novel cardioprotective drug.[21][22]
2Pharmacology
2.1. Absorption
Overall bioavailability of Berberine is quite low at 'less than 5%'[23][24] with 0.68% having been reported in rats.[25] Studies using 1,000-1,500mg Berberine by itself still appear to exert benefits after absorption, but enhancing absorption theoretically can reduce the dose of Berberine required to reach these effects.
Berberine appears to be subject to P-Glycoprotein mediated efflux from the intestines[26][27] and liver.[28] In the intestines, P-Glycoprotein is responsible for approximately 90% reduced transportation of Berberine[27] and Berberine has been further demonstrated to actually induce (increase) the activity of P-Glycoprotein transporters in the intestine[29][30] which have caused reduced absorption of other compounds subject to P-gp, such as Ciprofloxacin (demonstrated with 25-50mg/kg Berberine in rats; 4-8mg/kg human equivalent).[31]
Using an analogue that doesn't get subject to P-Glycoprotein (IMB-Y53) it is shown that increasing uptake contributes to further anti-diabetic effects[32] and pairing Berberine with compounds that are known to inhibit P-Glycoprotein (Milk Thistle,[33] Ketoconazole,[34] or d-α-tocopheryl polyethylene glycol 1000 succinate (TPGS)[25]) enhances the biological activity of Berberine.
Berberine has low rates of absorption when taken orally due to both being subject to P-Glycoprotein (ejects Berberine back into the intestines) and increasing the activity of P-Glycoprotein (augmenting its own ejection), but absorption is greatly increased when taken with P-Glycoprotein inhibitors such as Silymarin from Milk Thistle.
Absorption has also been enhanced with Sodium Caprate, a medium chain fatty acid that increases the size of tight junctions between intestinal cells (increasing paracellular permeability reversibly[35]) and appears to not be associated with adverse structural changes to the intestines when used with Berberine in vivo.[36][37] Sodium caprate is associated with improvements in AUC of Berberine by 28% (100mg/kg Berberine with or without 50mg/kg Sodium Caprate[37]) and appears to be further increased with 100mg/kg Sodium Caprate.[38] Enhanced absorption precedes greater post-absorptive effects, such as enhanced AMPK activation over 4 weeks (50mg/kg Sodium Caprate).[36]
Sodium Caprate, an ester of Capric Acid (Decanoic Acid; a constituent of milk fat at 2-3% and Coconut Oil at 10%) appears to enhance absorption via reversibly widening the gaps between intestinal cells and allowing passive diffusion. It is theoretical, but not yet demonstrated, that coingestion of Berberine with food sources of Capric acid could increase absorption of Berberine (and assuming 10% Capric acid content of Coconut oil, it is about 5.5g of Coconut oil for a 150lb human)
Due to low intestinal uptake rate, large doses (1g) are associated with constipation.[39] This constipative effect is also due to some properties of berberine in the colon, and can be useful to reducing watery diarrhea at 400mg, four 100mg doses.[40]
Low absorption may precede intestinal side-effects with high doses, due to large colonic levels
2.2. Distribution
Berberine binds to both Bovine and Human Serum Albumin relatively well and in a 1:1 ratio (indicating a single binding site) and has slightly higher affinity than does the related structure Palmatine;[41] Berberine may also bind to the β-Trp37 residue on Hemoglobin.[42]
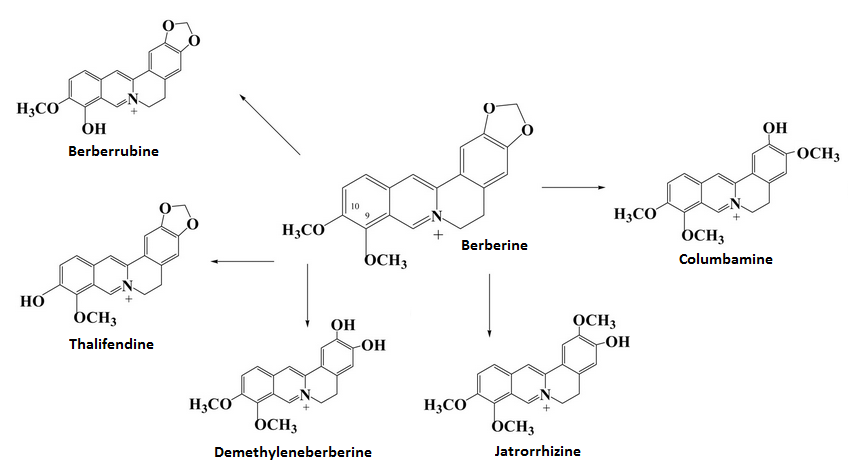
2.3. Metabolism
Berberine can have its structure metabolized into four possible metabolites known as Thalifendine, Jatrorrhizine, Berberrubine, and Demethyleneberberine; Berberrubine may passively isomerize between two molecules.[43][44] The metabolism of Berberine into its metabolites involves the enzymes CYP2D6 and CYP1A2 contributing 25.21% and 72.07% of metabolism into Thalifendine (CYP3A4 barely involved at 2.72%) with CYP3A4 as well as the former two being important to metabolism into Demethyleneberberine with a fairly even distrubion.[44]
One experiment with a molecular docking program suggested that CYP2E1, CYP2A6, and CYP2B6 were without any influence on Berberine and CYP2C19, CYP2C9 and CYP3A5 had very weak influence.[44] The lack of involvement of CYP2C9 and CYP2C19 were confirmed in HLMs in vitro.[44]
Berberrubine isomers exert the most potent AMPK activation and LDL receptor upregulation among the metabolites, but to a lesser extend than the parent compound Berberine;[44][43] this appears to also apply to the insulin receptor.[44]
Berberine can be possibly metabolized into four different metabolites, with all four metabolites both being active on the same mechanisms as Berberine but to a lesser potency
In rats, all four metabolites have been detected in serum following ingestion of 40mg/kg Berberine[45] and when measuring Berberine concentrations 3 hours after ingestion (rat liver) most Berberine appears to not be metabolized but a small increase in Thalifendine is noted relative to other metabolites.[46]
Berberine appears to be somewhat preserved in the parent form after oral administration
After incubation with intestinal bacteria for 7 days (human and rat), no visible metabolism of Berberine by intestinal bacteria was noted and the tested metabolites were similarly not metabolized further; it is thought that intestinal bacteria does not play a role in the metabolism of Berberine.[45]
2.4. Excretion
Orally ingested Berberine (chloride) at 900mg daily for 3 days was metabolized into three different urinary metabolites, with one (thought to be Jatrorrhizine-3-Sulfate) being the primary metabolite being excreted at 15-125 times more than the other two metabolites (Demethyleneberberine-2-sulfate and Thalifendine-10-sulfate, Berberrubine being undetectable in urine).[47] A later study noted that 900mg (3x300mg) for two days noted that Jatrorrhizine can be detected in the urine as a glucuronide (jatrorrhizine-3-O-β-D-glucuronide) as can Thalifendine (thalifendine-10-O-β-D-glucuronide), Berberrubine (berberrubine-9-O-β-D-glucuronide), and Demethyleneberberine (demethyleneberberine-2,3-di-O-β-D-glucuronide).[46]
One other metabolite has been found, columbamine-2-O-β-D-glucuronide;[46] both Jatrorrhizine and Columbamine can be found naturally occurring in Enantia chlorantha.[48]
Most urinary excretion of Berberine appears to be Jatrorrhizine, with all metabolites having at least once been detected in sulfated or glucuronidated form
2.5. Enzymatic Interactions
In vitro, Berberine appears to inhibit CYP3A4 with an IC50 of 48.9+/-9µM (16.4+/-3.0µg/mL).[49]
For human studies, three divided doses of 300mg Berberine (900mg total) confirmed CYP3A4 inhibitory potential as midazolam AUC was decreased 40%[50] and a few studies on Cyclosporin A (where serum levels increase due to CYP3A4 inhibition) have confirmed CYP3A4 relevance for humans.[51][52] This evidence is all in contrast to a previous rat study suggesting no effect of 100mg/kg bodyweight given an oral Carbamazepine.[29]
Many herbs that Berberine is derived from including Goldenseal[53][54] and Berberis Vulgaris[55] have been shown to inhibit CYP3A4 which is thought to be due to the Berberine content.
The inhibitory effect of Berberine on CYP3A4, an enzyme that metabolizes a fair bit of pharmaceuticals (the same one that St.John's Wort inhibits) appears to be relevant
Berberine has been shown in vitro to inhibit CYP1A2 (IC50 73.2+/-5.5uM; 24.6+/-1.8µg/mL) and CYP2D6 (7.4+/-0.36uM; 2.49+/-0.12µg/mL), suggesting possibly relevant inhibitory potential on CYP2D6.[49] A later study in humans confirmed biolologically relevant CYP2D6 inhibition by 900mg Berberine (in three divided doses of 300mg) and CYP2C9 was also found to be inhibited.[50] This study failed to note any significant inhibition of CYP1A2.[50]
Berberine has been noted to, in vitro, prevent the induction of a variety of CYP mRNAs and when administered to diabetic mice normalize CYP3A11, CYP4A10, and Cyp4A14[56] which are elevated during experimental diabetes.[57] Berberine also suppressed CYP2E1 in vivo.[56]
Various other enzymes appears to be inhibited following oral administration of Berberine to rats or in vitro
3Molecular Targets
3.1. AMPK
Adenosine Mono-phosphate Kinase (AMPK) is a nutrient sensor protein that is central to the actions of various anti-diabetic drugs (Metformin[58]), and appears to be a central lever point for the actions of Berberine. Berberine activates AMPK in a dose and time-dependent manner.[59][60] In investigating how Berberine induces AMPK (commonly associated with energy restriction or some hormetic agents), a possible mechanism is inhibition of complex I of the mitochondrial electron transport chain[17] which is also observed with the anti-diabetic drugs Metformin and Rosiglitazone (with similar efficacy to the latter, more than the former), resulting in 50% inhibition of respiration at 15umol/L Berberine.[17] Inhibition of this by overexpressing PGC-1α (genetically) can attenuate AMPK activation[61] as can the direct inhibitor Compound C (implicating both the mitochondria and AMPK itself in glucose uptake).[62] Inhibition of Protein Kinase C zeta (PKCζ; upstream of PKB) has also been found to inhibit Berberine-induced glucose uptake, where siRNA for PKCζ that reduced its activity by 50% inhibited 42+/-24% of Berberine-induced glucose uptake.[62]
This increase in AMPK appears to be biologically relevant as it has been found in vivo when rats are injected with 5mg/kg Berberine.[63]
AMPK is induced after Berberine administration, which has been observed in living systems. The regulation appears to be indirect. Mitochondrial uncoupling and a protein known as PKCζ both appear to be involved, but the exact mechanistic pathway it not fully established
AMPK activation by Berberine in HepG2 (liver) cells was found to inhibit both cholesterol and triglyceride synthesis with an IC50 value of 15ug/mL and reduced their respective IC50 values of 10.4 and 5.8 μg/ml.[64] These IC50 values are similar to those seen with LDL-C receptor upregulation and occured at similar time points,[65] suggesting that they are tied in to similar mechanistic roots. Although mediated by AMPK, inhibiting MAPK/ERK appears to attenuate the effects.[64]
Injections of Berberine into the brain also decrease Malonyl-CoA; which is increased when AMPK is inactivated (via ACC activity) and suggests that AMPK activation occurs in neural tissues.[63] This decrease in neural Malonyl-CoA may actually precede mitochondrial biogenesis in skeletal muscle.[66][67]
This activation of AMPK extends to adipocytes (fat cells), skeletal myocytes (skeletal muscle cell), and the liver; please refer to their respective sections for more information.
The indirect activation of AMPK appears to extend to a wide variety of tissues in the body
4Longevity and Life Extension
4.1. Telomeres
Binding of an agent to the 3' cap of telomeric DNA can interfere with the telomerase enzyme and give the appearance of a telomerase inhibitor, providing a novel mechanism of action for anti-cancer therapy with interest to life extension.[68] Compounds that tend to associate with this target are large aromatic compounds with a polar charge,[69][70] and Berberine has been found to bind to this target with a 1:1 stoichiometric ratio with relatively high affinity;[71] which has preceded Berberine derivatives being synthesized for higher affinity.[72][73] Binding specificities for Berberine can be read here.[74]
It is thought that telomerase inhibition of Berberine reaches 50% at 35mM concentration,[75] and expression of hTERT has been suppressed at 100ug/mL in SiHa and HeLa cancer cells.[76]
May have telomerase inhibitory potential; of interest mostly to cancer research but with some possible crossover into life extension. Currently no evidence to suggest how this influences lifespan in living models
5Neurology
5.1. Pharmacokinetics
Berberine appears to cross the blood-brain barrier and reach the brain parenchyma in a dose/time-dependent manner.[77]
Berberine can cross the blood brain barrier (BBB)
5.2. Adrenergic Neurotransmission
In regards to the Alpha-Adrenergic receptors (molecular targets of yohimbine), Berberine appears to have relatively more affinity for post-synaptic Alpha-1-Adrenergic receptors than presynaptic A2A receptors.[78] It appears to act as a partial agonist and potential competitive antagonist of these receptors, as assessed by platelets,[79] and appears to extend to its molecular class of Berbanes rather than being a unique property of Berberine.[80] Berberine has been found to interact with a binding site on the Beta-2-Adrenergic receptor.[81]
Appears to interact with adrenergic receptors; practical relevance unknown
Acute administration of Berberine at 5mg/kg injections can raise neural norepinephrine by 31% in mice.[82] Prolonging this treatment for 15 days maintains similar potency (29%), yet increasing the dose to 10mg/kg lessened the increase (12%, not statistically significant).[82] An increase in noradrenaline concent in both the hippocampus and frontal cortex (not striatum) have been noted following oral consumption of 20mg/kg bodyweight in mice, where noradrenaline was increased by 10.8% (hippocampus) and 26.1% (frontal cortex).[83]
Berberine is known to inhibit Monoamine oxidase enzymes with IC50 values of 126uM for MAO-A,[84]and 98.2-98.4uM for MAO-B.[85] These potencies are fairly low and likely not biologically relevant, although a metabolite of Berberine (Jatrorrhizine) has increased potencies on MAO-A and MAO-B with 6uM and 62uM, respectively.[84]
Has been shown to increase noradrenaline in mice brains following both injections and oral treatment, serum levels not yet measured
5.3. Serotoninergic Neurotransmission
Acute administration of Berberine at 5mg/kg injections can raise neural serotonin by 47% in mice, but chronic (15 day) dosing attenuates the increase to 19% but was preserved (53%) with 10mg/kg injections.[82] Oral ingestion of 20mg/kg in mice increased serotonin levels in the hippocampus (22.8%) and frontal cortex (23.6%) but not striatum.[83]
Has been shown to increase serotonin content in some areas of the brains of mice following oral ingestion; serum levels of serotonin following Berberine not yet known
5.4. Dopaminergic Neurotransmission
Berberine has been found to inhibit the tyrosine hydroxylase enzyme in PC12 cells in vitro.[86] This enzyme catalyzes the bioconversion of L-Tyrosine into the dopamine precursor L-DOPA.
Acute administration of Berberine at 5mg/kg injections can raise neural dopamine by 31% in mice, which increased to 53% over 15 days of administration; no further increase was noted at 10mg/kg injections over 15 days though (31%).[82] Ingestion of 20mg/kg Berberine in mice failed to significantly modify dopamine in the striatum, frontal cortex, or hippocampus.[83]
Has been shown to increase dopamine concentration in some areas of the brain following oral administration
5.5. Memory and Learning
Berberine has been noted to preserve Long-term potentiation (LTP) in perforant path-dentate gyrus synapses following 100mg/kg ingestion of Berberine over 11 weeks in rats.[87] LTP tends to be reduced in diabetes.[88][89] No alterations in cell count or apoptosis in the hippocampus were noted in this study,[87] despite apoptosis in the hippocampus being related to Diabetes-induced memory losses[90] and improvements in cognition as assessed by a spatial recognition memory in Y-maze as passive avoidance task were noted (not outperforming non-diabetic control, but outperforming diabetic control and 50mg/kg Berberine which was too low to be significant).[87] A reduced rate of diabetes-induced memory loss has been noted elsewhere with 25-100mgkg Berberine twice daily (total dose 50-200mg/kg) for 30 days in diabetic rats, which was credited to anti-oxidant effects secondary to AMPK activation (mimicked by both Metformin and high dose Vitamin Csupplementation).[91]
5.6. Depression
A mouse study using 5mg/kg Berberine (injection) noted time-dependent reduction in immobility time in a forced swim test, indicative of anti-depressive effects.[82] Berberine was also effective at reversing Reserpine-induced depression (which depletes catecholamines), and enhanced the anti-depressant effects of imipramine, tranylcypromine, fluoxetine, and venlafaxine but not trazodone or mianserine;[82] 10mg/kg fluoxetine (an SSRI) and 5mg/kg Berberine abolished the depressive effects in the forced swim test,[82] and the effect of Berberine (5mg/kg) is as effective as 10mg/kg Imipramine.[82] Anti-depressive effects have been noted following oral administration of 10-20mg/kg in mice, but 100mg/kg (human equivalent dose of 8mg/kg) was ineffective; it underperformed relative to 20mg/kg desipramine, but was synergistic with this one as well.[83]
Berberine appears to exert anti-depressive effects in mice, and appears to work synergistically with a wide variety of standard anti-depressant drugs; oddly, these effects have been observed at 20mg/kg in mice but not at 100mg/kg in mice (equivalent human doses of 1.6mg/kg and 8mg/kg, respectively) No human evidence, however
Oddly, these anti-depressant effects are abolished with injections of 750mg/kg L-Arginine (substrate of which nitric oxide is created) and Viagra (increases nitric oxide in the brain) and was enhanced by inhibitors of nNOS, an enzyme that creates nitric oxide in the brain;[82] this study is duplicated in Medline.[92]
Anti-depressant effects can also be traced back to Sigma receptors, where activation of Sigma receptors σ1 enhances anti-depressive effects and antagonism abolishes the anti-depressive effects.[82][92] Berberine is known as a positive Sigma1 receptor modulator, which is considered a relatively new class of therapeutic options for depression.[93] Sigma receptors are novel intracellular receptors (expressed on the endoplasmic reticulum (ER)[94]) and their activation may modulate glutamingeric signalling such as NMDA.[95] They can regulate calcium signalling at both the level of the ER as well as cytoplasm,[96] and a known naturally occurring ligand for S1 receptors is the hallucinogenic N,N-dimethyltryptamine[97] (not to be confused with Hordenine, N,N-dimethyltyramine).
The mechanism of action for Berberine and anti-depression may be via acting as a positive modulator of Sigma-1 receptors (molecular target of the hallucinogen DMT), and enhancing signalling via this receptor; this is likely to occur at rat oral doses of 20mg/kg (preliminary evidence) which correlates to a human oral dose of 1.6mg/kg bodyweight yet may not occur at five-fold the dose (8mg/kg)
5.7. Sedation
Although injections of 2-5mg/kg Berberine fail to alter locomotion, 20mg/kg injections can reduce locomotion in mice[82] (same dose not effective after oral administration[83]). This high dose injection also augmented phenobarbitol-induced sleep time, with lower doses ineffective.[82]
May have sedative properties at higher doses
5.8. Analgesia
A study assessing the analgesic effects of berberine noted that chronic (7 day) treatment of 10mg/kg injections reduced nocioreception in a tail-flick test, with the potency being greater than 10mg/kg Imipramine.[82]
5.9. Alzheimer's Disease
Berberine has been found to have inhibitory potential against Acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) with IC50 values of 0.37-0.58uM for AChE[98][99][100] and 3.44uM for BChE.[98] A PANTHER analysis[101] (computer program) noted that another potential targets include Amyloid-β (A4) precursor protein[102] but that assigning Berberine's targets into known pathways yielded 11 targets in the Alzheimer disease-amyloid secretase pathway and 17 targets in the Alzheimer disease-presenilin pathway. [101]
In vitro, Berberine can reduce β-Amyloid levels (APPNL-H4 cells) by 47.1+/-11.5% and 49.1+/-13.6% (Aβ40 and Aβ42, respectively) at 5uM with no further benefit at 50uM[102] possibly secondary to stimulating α-secretory activity (311.9+/-7.9% of control) and downregulating β-secretase (55.5+/-11.1%).[102] The downregulation of β-secretase appears to be secondary to ERK1/2 activation by Berberine.[103]
May beneficially alter the content of β-Amyloid levels in isolated neurons, which is thought to be therapeutic for Alzheimer's Disease. May also have pro-cholinergic effects via enzyme inhibition (also thought to be therapeutic) at a potency that might be relevant in vivo
One rat study using Phellodendron amurense at 100-200mg/kg but with another group injected with 20mg/kg isolated Berberine for 14 days prior to scopolamine injections (anti-cholinergic toxin) was able to attenuate deficits to learning, with an efficacy lesser than 0.2mg/kg tacrine used as active control.[104]
5.10. Neuroprotection
Pretreatment of Berberine in the range of 10nM to 1µM caused dose-dependent cell preservation (up to 65% cell preservation at 1µM) in response to a research oxidative toxin (CoCl2)[105] and survival of neurons has been noted when Berberine injections are given prior to ischemia/reperfusion surgery in rats.[106] Berberine does not cause any cytotoxicity in neurons up to 10µM.[105]
Conversely, when PC12 cells (Pheochromocytoma, used to assess survival and differentiation in vitro) are incubated with 6-OHDA (a pro-oxidative metabolite of dopamine that may induce cell death) and subsequently 10-50µM Berberine, Berberine augments 6-OHDA induced neurotoxicity in a dose-dependent manner.[107] The concept of Berberine augmenting PC12 toxicity in the presence of 6-OHDA has been reported elsewhere,[86] and this has been found in response to CoCl2only when Berberine was applied after the toxin.[105] Pretreatment of a toxin followed by Berberine appears to enhance cytotoxicity, which may be due to Berberine's interaction with the mitochondrial since it interfere with mitochondrial complex I (to activate AMPK[17]) but excessive inhibition of this complex, as is seen with the MPP+ toxin[108] is neurotoxic.
There appears to be a protective effect of Berberine when it precedes toxins (preventative), but Berberine applied after the toxin (therapy) appears to have limited evidence to suggest potentiation of toxicity. This may be due to berberine's main mechanism (inhibition of mitochondrial complex I) inhernetly being hormetic and hormesis not being the smartest thing in instances of cellular damage as is seen with pretreatment of a toxin
When rats were lesioned with 6-OHDA (metabolite of dopamine used in research to mimic Parkinson's Disease lesions) and then subsequently orally treated with 5 or 30mg/kg Berberine (injections) for 21 days, 30mg/kg but not 5mg/kg was associated with less surviving neurons in the substantia Nigra.[107] This mechanism appears to be common to isoquinoline derivatives such as tetrahydropapaveroline, salsolinol, and TIQs.[109]
This adverse effect of Berberine therapy appears to occur in living models
5.11. Addiction and Dependence
One study in morphine dependent mice noted that Berberine (50mg/kg injections) was able to partially normalize the reductions in BDNF mRNA, CRF, and TH expression which predisposed morphine-dependent mice to anxiety and depression.[110] The dose of 50mg/kg Berberine was able to normalize anxiety and depression as was 10mg/kg Fluoxetine, with 10-20mg/kg showing a trend to benefit (not statistically significant).[110]
May attenuate withdrawal from Morphine, may require high doses to do so; no human evidence nor oral consumption evidence currently
6Cardiovascular Health
6.1. Cardiac Tissue
Ischemia-Reperfusion injury in cardiac tissue may be alleviated by Berberine. Rats fed 100mg/kg Berberine daily for 14 days prior to insult (both in vitro testing of cardiac tissue and in vivo testing conducted) Berberine pretreated hearts were associated with preservation of LVDP (75%), LVEDP (29%), +dp/dtma (75%), and −dp/dtmin (69%) without any influence on these parameters per se(without IR injury).[111] Protective effects were noted in vivo as well (LVDP by 9-10% and LVEDP by 40-45%) alongside reduced infarct size and protection from arrythmia; these protective effects are thought to be related to AMPK regulation, with a reduction in AMPK phosphorylation noted in the Berberine IR group.[111] These protective effects have been noted elsewhere with Berberine in diabetic rats[112] and may be related to the structural class rather than being novel to Berberine, as both Coptisine[113] and Palmatine[114] have similar protective effects.
May protect cardiac tissue from Ischemia-Reperfusion (oxidative) injury via AMPK, but the regulatory effects on AMPK are atypical of Berberine's other AMPK-related actions
In regards to cardiac fibrosis, Berberine has once been noted to form G-quadruplexes with the rat relaxin-1 gene promoter region, which attenuated STAT3 downregulation of Relaxin expression (STAT3 is a negative regulator of Relaxin[115]) to 55% of control (20uM) and indirectly promoted mRNA and protein content of Relaxin.[116] Due to Relaxin's role in preventing cardiac fibrosis,[117]Berberine administration (injections) resulted in less fibroblast activation, collagen synthesis, and extent of cardiac fibrosis.[116] Berberine administration appears to promote relaxin in a dose-dependent manner, with 100uM injections increasing Relaxin more than 2.5-fold relative to baseline, and did not inherently influence STAT3 expression nor did it influence nuclear translocation of STAT3 following activation.[116]
May have potential to reduce cardiac fibrosis, but limited evidence with no studies assessing oral Berberine ingestion nor comparison to reference drugs (to assess potency)
Berberine has been shown to have Muscarinic (M2) acetylcholine receptor agonist activities, which has been noted in cultured rat cardiomyocytes where antagonists of the Muscarinic receptor but not beta-blockers abolished the benefits on heart tissue contractility.[118]
These M2 agonist properties may underlie a reduction in heart damage that has been observed, where 150-300mg/kg Berberine was able to attenuate the adverse cardiac effects of a high carbohydrate diet in diabetic rats[119] and may explain how 1.2-2g Berberine daily in persons with cardiomyopathy (in addition to standard therapy) increases Left Ventrical Ejection Fraction and improves quality of life more than placebo.[120]
Berberine also exerts protective effects on the heart after acute injury.[121]
6.2. Triglycerides
30mg/kg Berberine has been found to preserve the protein content of the LDL receptor during periods of inflammation in rats,[122] and may also induce its upregulation via JNK;[123] this is most likely relevant in humans as LDL cholesterol has been shown to be reduced by 25% in hypercholesterolemic persons over 3 months of Berberine ingestion.[65]
Beyond modifying the expression of the receptor, activating ERK can also preserve the constitution of the receptor and stabilize it; leading to a prolongation of the time the LDL receptor can uptake LDL-C into liver cells.[124]
One study investigating proprotein convertase subtilisin/kexin type 9 (PCSK9; also known as NARC-1 or proprotein convertase 9), an enzyme that degrades the LDL receptor and increases serum LDL as a consequence,[125] noted that Berberine at an oral dose of 10-30mg/kg in rats attenuated the spike in PCSK9 associated with inflammation which was associated with somewhat normalized lipid parameters in blood.[122] PCSK9 is a molecular target connecting inflammation to LDL-C since its expression is positively correlated with inflammation.[126]
The LDL receptor (which takes up LDL from the blood and contributes to an LDL lowering effect) can have its degradation attenuated and its synthesis increased by Berberine by two separate mechanisms; this may underlie the recorded reductions of LDL following Berberine administration to humans which has reached up to 25% in 3 months in people with high cholesterol
Berberine may inhibit lipid synthesis, secondary to the activation of AMPK seen as the main mechanism of action.[127]
Triglycerides have been noted to be reduced 16% following 1g of Berberine ingestion for 4 weeks[2]and 1500mg daily for 12 weeks in otherwise healthy obese persona has noted a reduction of 23%.[128] A placebo-controlled challenge-rechallenge in people with high cholesterol but who were otherwise at low risk for cardiovascular disease also found a statistically significant lowering of triglycerides during the second rechallenge (but not the first) along with a lowering of LDL-C and raising of HDL-C during both phases when compared to placebo; anthropometric measurements and blood sugar were not effected by berberine, although these patients did not have high blood sugar at the start of the trial.[129] A meta-analysis conducted on diabetics (with concurrently high triglycerides) has noted that the reduction of triglycerides averaged 0.48mmol/L with a CI 0.39-0.57, reaching statistical significance.[130]
AMPK activation is a mechanism by which triglycerides can be reduced, which has been reported in humans and appears to be reliable
One meta-analysis has been conducted on diabetic persons that also measured lipid parameters as endpoints, and this meta-analysis concluded significant reductions in Triglycerides (0.48mmol/L reduction; CI 0.39-0.57), Total Cholesterol (0.58mmol/L; CI 0.14-1.02), and LDL-C (0.58mmol/L reduction; CI 0.39-0.78) with an increase in HDL-C (0.07mmol/L increase; CI 0.04-0.10) in these diabetic patients;[130] specifics of the meta-analysis can be reviewed in the Glucose Interventions section.
Appears to have beneficial effects on lipid parameters in Diabetics
6.3. Blood Flow
Berberine is a known vasorelaxant in animals[131][132] and has been used with success in humans.[133]
Appears to have vasorelaxing properties
It is known that berberine works on the endothelium itself and underlying smooth muscle (mostly the former)[134] and it is thought that berberine may work via ACE enzyme inhibition of the NO-cGMP axis[135] or through α1-adrenergic receptor blocking.[136][78] One study has noted that berberine at 25-200µM is able to inhibit ephedrine and histamine induced aortic contractions in a reversible manner yet failed to inhibit contractions from high potassium or caffeine.[137]
6.4. Platelets
Anti-platelet functions have also been noted with berberine, including inhibition of thromboxane synthesis[138] and increased thrombolysis (breaking of clots).[139][133] Other possible mechanisms of action on its anti-thrombotic effects are alpha(2)adrenoceptor agonism on platlets[79] and an inhibitory effect on calcium influx.[140]
6.5. Atherosclerosis
Berberine (10-50uM) can suppress the activity of a protein known as AEBP1, which prevents the uptake of oxidized LDL by macrophages in a dose-dependent manner and attenuates formation of foam cells (artherogenic deposites of macrophages).[141] Suppressing AEBP1 leads to less expression of the scavenging receptors LOX-1 and CD36 and less influx of oxidized LDL (which promotes foam cell formation); CD68 was unaffected by Berberine.[141] Secondary to preventing oxLDL influx, secretion of adhesion factors (proteins secreted from immune cells to promote adhesion) ICAM-1 and VCAM-1 are both suppressed.[142]
One study on rats that lacked expression of AMPK noted that the beneficial effects of Berberine on artherosclerosis (including reduced cardiac lesions and oxidative stress) was mostly abolished, suggesting AMPK is critical for cardioprotective effects of Berberine.[143]
7Fat Mass and Obesity
7.1. Mechanisms (Fat Cell Regulatory)
In preadipocytes incubated with 1-10uM Berberine for 3 days during differentiation noted that there was an increase in proliferation (not dose dependent, also seen in mouse adipocytes[144] and 3T3-L1[145]) and suppression of differentiation (dose dependent) up to half suppression at 10uM; post-differentiation, the protein content for LPL, C/EBPα, and PPARγ2 was significantly reduced by more than half and leptin and adiponectin protein content and secretion was significantly reduced.[146] In 3T3-L1 adipocytes, an inhibition of both proliferation and differentiation has been noted at 1.25-5uM, and replicated a suppression of C/EBPα and all three PPAR subsets (α, β/δ, and γ).[145]
Although AMPK is known as the main mechanism of Berberine, it appears that Berberine (3uM) activates an unfolded protein response independent of endoplasmic reticulum stress (vast majority of the time, it is directly linked[147]) which induces activity of CHOP; inhibiting CHOP appears to abolish the reductions in PPARγ and C/EBPα.[148]
In preadipocytes, Berberine appears to suppress differentiation and has mixed effects on proliferation; both studies suggest a reduction in the pro-adipogenic receptor PPARγ (which, oddly, is anti-diabetic when activated)
In mature adipocytes, a suppression of PPARγ is still noted at 1.25-10uM concentration at both the mRNA and protein level; incubation with 10uM Berberine and PPARγ activators (troglitazone and rosiglitazone) demonstrated that PPARγ in inhibitory and reduced the effects by 70-80%.[145] This inhibitory potential extends to PPARα, but to a lesser degree (40-60%), and Berberine was said to notbe a ligand of the PPAR recepetors.[145] This may be related to the aformentioned activation of an unfolding protein response, which induces CHOP activity and suppresses PPARγ and C/EBPα activity; this was noted to also occur in mature adipocytes.[148] The influence of the WNT/β-Catenin pathway has been ruled out.[149]
When cells are taken from diabetic rats (diabetes induced, fed a high carbohydrate diet, then white adipose tissue excised), appeared to reduce TNF-α concentration (-39%) and increase content of all PPAR subsets with the induction of PPARδ and PPARα similar to Fenofibrate and PPARγ similar to Rosiglitazone.[150] These benefits were also seen in another group of rats fed Berberine at 75-300mg/kg for 16 weeks, where both 150 and 300mg/kg outright normalized serum and adipose Lipoprotein Lipase (LPL), serum free fatty acids and TNF-α, and expression of all PPAR subsets and Cyclin T1 (potency was similar to the additive benefits of 100mg/kg fenofibrate and 4mg/kg rosiglitazone).[150]
In mature non-diabetic adipocytes, appears to suppress the activity of PPARy (a protein that is pro-adipogenic). In diabetic adipocytes (confirmed in vivo), Berberine can normalize expression of PPARs that is normally suppressed
Berberine may have anti-lipolytic effects.[151] This study noted that while Berberine did not influence baseline cAMP levels, stimulation of cAMP via PDE inhibitors was attenuated (not abolished) by Berberine; Berberine lessened the inhibitory effects of other compounds, without inherently inhibiting the enzyme action independently of AMPK.[151] This anti-lipoytic effect has been noted elsewhere in 3T3-LI adipocytes where lipolysis was stimulated by IBMX, forskolin and 8-bromo-cAMP.[152]
Berberine may induce HSL activity (a pro-lipolytic effect) via an AMPK independent manner.[151]
Theoretically possible that Berberine can reduce the fat burning effects of PDE inhibitors (luteolin, resveratrol) and forskolin (active ingredient in coleus forskohlii)
7.2. Mechanisms (Glucose Related)
In regards to adiponectin, an adipokine (signalling molecule derived from fat cells) that plays a positive role in insulin sensitivity (is secreted, and then acts on tissue via its receptors to activate AMPK[153]) and is reduced in diabetics, particularly the high activity structure.[154] Adiponectin is found in three structural forms (trimer, hexamer, and high molecular weight) with the latter being the most related to insulin sensitivity;[155] Berberine (2-4uM) acts through AMPK activation, particularly the AMPKα1 subset, to increase the percent of adiponectin in high activity structure; a process known as adiponectin multimerization.[156] This was also noted with AICAR, a research drug used to activate AMPK, suggesting a general effect that is not unique to Berberine;[156] this intricate loop (Adiponectin activating AMPK which promotes high-activity adiponectin) is a mechanism of adiponectin self-regulation.[157]
It should be noted that studies using Berberine in predifferentiated adipocytes noted less secretion of adiponectin which was the natural consequence of suppressing differentiation.[146] This was also replicated in the aforementioned study on enhancing adiponectin function, with both phenomena occurring at similar concentrations.[156]
Appears to enhance the activity of adiponectin, but practical relevance of this mechanism for insulin sensitivity is unknown
Berberine is known to enhance glucose uptake into fat cells, and at 25uM concentration it is equally potent as 15uM 2,4-thiazolidinedione (TZD, an anti-diabetic drug, by 3.3 fold) and slightly outperformed both arecoline (3.2 fold) and vanillic acid (2.9 fold), natural products.[158] This study also noted that Berberine acted synergistically with both TZD and Metformin.[158] Berberine has also been shown to be more effective at enhancing glucose uptake than polysaccharides from Astragalus Membranaceus.[159]
Although AMPK activity increses by Berberine is known to increase glucose uptake into adipocytes,[160] Berberine seems to act independently of AMPK to increase glucose uptake by 5-fold in L929 fibroblast cells that only express GLUT1 transporters; Berberine was found to increase the activity of GLUT1 (a normally low-active glucose transporter, with GLUT4 being the major one) via a partially p38 MAPK and ERK dependent pathway.[161] This increase in GLUT1 activity has been noted elsewhere in adipocytes (3T3-L1), although attributed as secondary to AMPK activation.[162]
Berberine may also inhibit the PTP1B enzyme and promote glucose uptake into adipocytes (and myocytes) by preserving the actions of insulin. At concentrations of 1.25-2.5uM Berberine, insulin receptor phosphorylation is increased without affecting protein content.[160] The IC50 of Berberine on PTP1B appears to be 156.9nM and a Ki value of 91.3nM, remarkably potent.[163]
Berberine has been found to partly normalize the decrease of glucose uptake induce by palmatate (a fatty acid), which is through anti-inflammatory effects in inhibiting increased activity of IKKβ and NF-kB; which subsequently increase IRS-1 and reduce glucose uptake via the insulin receptor.[164] This anti-inflammatory effect has been noted elsewhere when measuring cytokines,[165] and fatty-acid induced insulin resistance has also been replicated elsewhere related to NF-kB.[166]
Multiple mechanisms of increasing glucose uptake into fat cells, despite inhibting their proliferation. Augments insulin-dependent glucose uptake (PTP1B), insulin-indepenendent glucose uptake (AMPK), increasing the affinity of low-activity glucose transporters (GLUT1), and attenuating insulin resistant effects
7.3. Interventions
One study in persons with newly diagnosed metabolic syndrome noted that 300mg Berberine thrice a day (900mg total) for 12 weeks was associated with a significant reduction of BMI from 31.5+/-3.6 to 27.4+/-2.4 (average 13% decrease) with a significant decrease in waist circumference by 5.5%; lean mass and fat mass were not measured.[146] Otherwise healthy but obese persons taking 500mg Berberine thrice daily (1500mg total) for 12 weeks without adjustments to exercise noted reduction in body weight of approximately 5lbs (2.3% body weight; 3.6% body fat); food intake was not changed overall, but two subjects reported a decrease in appetite.[128]
Another trial in humans with nonalcoholic fatty liver disease who were given either lifestyle interventions alone, or lifestyle interventions plus 15mg pioglitazone daily or 0.5mg berberine thrice daily for 16 weeks found a significant reduction in BMI of 1.51 in the berberine group compared to a 0.72 reduction in BMI with lifestyle interventions alone; pioglitazone had a reduction similar to lifestyle interventions alone.[167]
Not too much evidence in humans, but some studies confirm weight loss with one suggesting a very slight preference for fat loss based on percentages; seemingly more potent in unhealthy persons, no studies in persons of normal weight yet.
8Interactions with Glucose Metabolism
Note: For a comprehensive review of how Berberine can interact with glucose metabolism, the sections on the Liver and the Pancreas (under 'Interactions with Organ Systems') should be reviewed, as should subsections in both the section on Fat Mass and Skeletal Muscle Metabolism that mention glucose. These four sections contribute to the potent anti-diabetic effect of Berberine
8.1. Absorption
Berberine appears to weakly suppress glucose uptake acutely,[23] with 72 hours of incubation suppressing glucose uptake to a statistically insignificant degree in vitro.[168]
Sucrase is inhibited in a concentration-dependent manner with an IC50 of 1.83mg/L (fairly low potency)[23] ot 0.28mg/mL.[169] The sucrose-isomaltase (SI) enzyme complex appears to have its mRNA increased in the state of diabetes, which is reduced (to up to 62% that of control rats) at 100-200mg/kg Berberine for 35 days, this affected non-diabetic rats as well.[170] Another study using 125mg/kg Berberine for 33 days noted that, in response to an oral sucrose tolerance test, that Berberine resulted in 43% less AUC for glucose in serum (less efficacy than 20mg/kg Arcabose as active control) which correlated with less sucrase activity in all parts of the intestines.[169] 100mg/kg Berberine for 4 weeks has been reported to have similar effects elsewhere.[171]
Maltase appears to be inhibited but not in a dose-dependent manner,[23] with another study suggesting the alterations of Maltase activity seen in diabetic rats (1.45-2.56 fold increase) being normalized with 35 days of supplementation of Berberine (100-200mg/kg)[170] and mostly normalized in all areas of the intestines after 125mg/kg for 33 days.[169] There does not actually appear to be any direct inhibition of active maltase enzymes up to 50uM of Berberine (somewhat contested, another study suggests an IC50 of 0.11mg/mL[169]), but 5 days of exposure to this concentration reduces activity by 48% in vitro.[170]
Lactase (mediates digestion of lactose) is also increased in the state of diabetes and attenuated, but not normalized, following ingestion of 125mg/kg Berberine for 33 days in rats.[169]
In regards to alpha-amylase (mediates starch digestion), Berberine has been tested in an in vitroinhibition test for fungal amylase noted dose-dependent growth inhibition with Ki values similar to Chlorogenic Acid and Caffeic acid and suggested non-competitive inhibition of the enzyme itself.[172]
The downregulation of enzyme activity (Maltase and SI complex) appears to be partly PKA dependent, and inhibiting PKA with the inhibitor H89 attenuates (but does not abolish) these effects.[170]
Direct inhibition of carbohydrate digestive enzymes appears to be either weak or non-existent. Prolonged treatment of Berberine may cause a reduction in synthesis of the enzymes that mediate sugar absorption (lactose, sucrose, and maltose) with possible but currently unexplored direction inhibition of the enzyme mediating starch absorption
Studies that compare the potency of Berberine to the reference drug, Arcabose, suggest that it is slightly (statistically significant) less potent
8.2. Interventions
The hypoglycemic effect of Berberine was first discovered in 1988 when a hypoglycemic effect was accidentally noted in diabetic patients when Berberine was given for anti-diarrheal effects.[173]
One Meta-Analysis has been conducted on Berberine has been conducted as it pertains to Type II Diabetes.[130] This Meta-Analysis noted 14 trials (all originating from China) including 1068 patients between the years of 2007-2011 and noted that Berberine at 0.5-1.5g daily paired with lifestyle intervention over 12 weeks was associated with improvements in Fasted (0.87mmol/L reduction; CI 0.54-1.20) and Postprandial (1.72mmol/L reduction; CI 1.11-2.32) blood glucose and HbA1c (0.72% reduction; CI 0.47-0.97%) with improvements in lipid metabolism and a reduction in Fasting Insulin Levels (0.5mU/L; CI 0.03-0.96).[130]
7 Trials (of 448 patients) used comparative assessment against oral hypoglycemic agents and, although a meta-analysis could not be performed due to heterogeneity of data, that there did not appear to be any significant differences when Berberine was compared against Metformin, glipizide, or rosiglitazone.[130] In 4 out of 6 trials that used Berbering as adjuvant treatment alongside oral hypoglycemics, additive benefits were found to be significant with Fasting (0.59mmol/L reduction; CI 0.35-0.83) and Postprandial (1.05mmol/L reduction; CI 0.48-1.62) blood glucose as well as HbA1c (0.53% reduction; CI 0.11-0.95%) dropping more in combination therapy than with oral hypoglycemic drugs alone.[130]
Methodology of the included studies was deemed subpar (Jadad score less than 3) but there did not appear to be risk of bias as assessed by funnel plot (although due to less than 10 studies being used, funnel plot may not have been as accurate as desired[174]).[130] This meta-analysis excluded three studies (none of which are indexed online) due to differences at baseline or uncertainty in randomization.[130]
Other trials on Berberine note that 0.3g thrice a day (900mg total) for 12 weeks in 37 persons with newly diagnosed metabolic syndrome noted significant reductions in blood glucose (17%), HbA1c (15%), fasting insulin (26%), and insulin sensitivity assessed by HOMA-R (41%).[146] Type II Diabetics given 1g Berberine for a month experiencing 20% and 26% reductions in fasting and postprandial blood glucose alongside a 12% reduction in HbA1c, but only a slight trend to improvement in insulin sensivity.[39] 1g Berberine over 2 months reducing fasting blood glucose (25.9%), HbA1c (18.1%), and triglycerides (17.6%).[175] Another trial in humans with nonalcoholic fatty liver disease who were given either lifestyle interventions alone, or lifestyle interventions plus 15mg pioglitazone daily or 0.5mg berberine thrice daily for 16 weeks found no difference between the 3 groups in HbA1C, but did find improved HOMA-IR scores in the berberine plus lifestyle interventions group compared to lifestyle interventions alone, with no difference versus lifestlye interventions plus pioglitazone.[167] Area under the glucose curve after an oral glucose tolerance test was also reduced in the berberine group compared to lifestyle interventions alone (with again no difference versus pioglitazone), primarily due to stronger glucose lowering at the 120 and 180 minute mark.[167] Similar improvements were found in people with the metabolic syndrome, where 0.5g of berberine three times a day for three months resulted in improved insulin sensitivity as measured by the insulinogenic and Matsuda indices, glucose AUC glucose, and insulin AUC versus placebo.[176]
Comparative studies using Berberine note that 1g daily find that it is equally effective at improving measured parameters (usually fasting blood glucose, insulin, HbA1c, and triglycerides) when compared to Metformin[177][175] and Rosiglitazone[175] when they are used within the standard dosage range of 1.5g (Metformin) or 4mg (Rosiglitazone).
Appears to be beneficial for blood glucose control in diabetic persons, and the potency of 0.5-1.5g daily does not appear to be significantly different than standard anti-diabetic pharmaceuticals and may be additive
9Skeletal Muscle and Physical Performance
9.1. Mechanisms (Glucose Related)
Berberine has been shown to stimulate glucose uptake into skeletal muscle[178] partially via AMPK mediated effects.[179]
AMPK activation can increase mitochondrial biogenesis in skeletal muscle cells, which Berberine has been shown to do; the inactivity of Berberine in cells lacking SIRT-1 (a required intermediate) has been established.[180]
Appears to increase glucose uptake and mitochondrial biogenesis in muscle cells via AMPK
The inhibition of PTP1B, which promotes insulin receptor signalling with an IC50 of 156.9nM,[163] also appears to occur in muscle cells.[160]
Berberine appears to induce glucose uptake into muscle cells by itself whether the muscle cell is insulin resistant[181] or insulin sensitive,[62] but a synergistic interaction between insulin and Berberine only exists when the muscle cell is insulin resistant, with insulin sensitive cells being barely additive (additive effects not statistically significant, there appears to be crossover in the mechanisms).[181][62]
The upregulation of the insulin receptor protein content (amount of insulin receptor expressed on the cell surface) appears to extend to L6 rat myocytes at 7.5uM to 2.5-fold that of control, with significant but lesser benefits noted at 2.5uM.[182] This was due to increasing transcription of the receptor at the genomic level, and is PKC dependent, which Berberine appears to dose-dependently activate.[182]
Can enhance both the expression of the insulin receptor and improve its signalling (via PTP1B inhibition), fairly potent at the latter in a nanomolar range. PKC and its subset PKCζ (zeta) appear critical in the potentiation and preservation of insulin signalling as well
9.2. Mechanisms (Proliferation and Differentiation)
Berberine (5mg/kg injections) has been found to induce muscle protein atrophy in mice by stimulating breakdown and inhibiting synthesis, an increase in atrogin-1 appeared to mediated these effects, and was independent of Akt/PI3K and FoxO alterations but found to be related to AMPK activation[61] which is known to increase Atrogin-1.[183] The induction of muscle protein breakdown was mediated by the ubiquitin-proteasome system and the attenuating of muscle protein synthesis by reducing content of eIF3-f; both of these effects were abolished by Atrogin-1 siRNAs, suggesting that this protein (induced by Berberine) is causative.[61]
The increase of Atrogin-1 may also be partly mediated by mitochondrial stress, as overexpression of PGC-1a prevents Berberine-induced Atrogin-1 release.[61] Berberine has been found to interfere with complex I of the mitochondria, and augment protein uncoupling (UCP2,3)[63][17] and this interference is the same thing that underlies AMPK activation from Berberine (as well as Metformin).[17][58] When the Myotubes were overexpressing PGC-1α, the decrease in myocyte diameter was abolished as was the increase in AMPK activation.
AMPK activation appears to lead to suppressed Myocyte growth, which is mediated by Atrogin-1 induction; overexpression of PGC-1α prevents this, but may abolish the AMPK induction. Abolishing Atrogin-1 reverses suppression into hypertrophy. Limited evidence all around
10Skeleton and Bone Metabolism
10.1. Calcitriol
A study in healthy obese subjects given 500mg Berberine thrice a day (1500mg total) for 12 weeks noted a trend (p=0.11) to increase serum calcitriol was seen in all subjects by 59.5%; this increase in the hormonally active form of Vitamin D was not tested in the concomitant rat study and the influence of seasonal changes cannot be ruled out.[128]
10.2. Joint Inflammation
In vitro with synoviocytes (proliferation of which is involved with pathology of Rheumatoid Arthritis), Berberine was able to inhibit cell proliferation at G0/G1 phase (thought to be from reducing mitochondrial membrane potential).[184] In chondrocytes, Berberine was able to attenuate MMP concentrations (seen as being involved in osteoarthritis[185]) and increase TIMP-1 at 25-100uM, which worked against IL-1b actions and exerted an anti-osteoarthritic effect; 50uM showed almost normalization of these levels to control values.[186]
Shows mechanisms that could aid both Rheumatoid and Osteoarthritis, moderately potent
In a rat model of adjuvant-induced Arthritis, 10mg/kg Berberine (injected) daily 9 days was able to attenuate the paw edema (marker of disease progression) in mice while a shorter supplement time frame of 3 days exerted nonsignificant benefit and irregular injections actually exacerbated paw edema.[187] An acute injection of 50-100uM Berberine into a rat knee three hours prior to inflammatory insult was able to abolish the effects of IL-1b at 100uM Berberine.[186]
Highly potent anti-inflammatory effects, but the animal models use injections; not known how oral ingestion affects joint health
11Inflammation and Immunology
11.1. Inflammation (General)
Oral ingestion of Berberine for 4 weeks can dose-dependently reduce the serum rise of 8-isoprostane in response to the pro-inflammatory LPS, with oral ingestion of 30mg/kg effectively abolishing the LPS-induced rise in 8-isoprostane.[122] The LPS-induced increase of TNF-α (53-86%), IFN-γ (74-88%), and IL-1α (68-93%) was also attenuated with 10-30mg/kg ingestion of Berberine.[122]An attenuation of these proinflammatory biomarkers has also been noted in response to dextran sulfate sodium (research chemical to induce colitis)[188] and in response to TNBS-induced colitis, where the degree of cytokine reductions following 20mg/kg reached 100% (TNF-α), 78% (IL-1β), and 98% (IL-6) while the levels of IL-10 (reduced to 11% in colitis control) had the reductions attenuated to 53%.[189]
Chronic ingestion of Berberine appears to be associated with less inflammation in response to pro-inflammatory stimuli.
When looking at the interaction of Berberine and Cyclooxygenase enzymes, Berberine does not interact with COX1 or COX2 at concentrations up to 100uM.[190]
11.2. Macrophages
Acutely, Berberine incubation with RAW264.7 macrophages that are stimulated with LPS (proinflammatory agent) appears to have an IC50 value in inhibiting nitric oxide release of greater than 30uM (weak effect).[191]
Has not yet been demonstrated to have effective anti-inflammatory effects in acute studies on macrophages
Berberine (10-50uM; concentrations below 75uM are nontoxic[192]) can suppress the activity of a protein known as AEBP1, which prevents the uptake of oxidized LDL by macrophages in a dose-dependent manner and attenuates formation of foam cells (artherogenic deposites of macrophages).[141] Suppressing AEBP1 leads to less expression of the scavenging receptors LOX-1 and CD36 and less influx of oxidized LDL (which promotes foam cell formation); CD68 was unaffected by Berberine.[141] Suppression of LOX-1 and another scavenger receptor, SR-BI, were noted to nearly control levels at 5-10mg/L Berberine over 24 hours incubation with no effect on ABCA1.[193]
This inhibition of oLDL uptake may also prevent other consequences of oLDL uptake, such as MMP9 secretion and NF-kB activation in macrophages.[192]
May prevent macrophages from becoming foam cells, which is a consequence when they absorb oxidized LDL (an acutely protective effect, but over time makes the foam cells themselves become plaque in arteries)
11.3. T-Cells
Berberine at 10-20mcg/mL concentrations in vitro appears to slightly enhance T-cell proliferation in response to antigens, while concentrations above that show dose-dependent immunosuppression.[187]
Berberine appears to be an selective inhibitor of JAK3 with 20-fold more affinity for JAK3 than JAK2 (with no apparent affinity for JAK1),[194] the JAK3/STAT pathway being one that induces a fair bit of inflammation from cytokines in models of arthritis and mediates immunology (inhibition of which is a novel class of immunosuppressants).[195][196] Berberine can inhibit IL-2 induced signalling via JAK3 with an IC50 of 3.78µM and the IC50 on JAK2 was 80µM, and in vitro at 3µM Berberine nuclear activity of JAK3 (via STAT5) is mostly undetectable.[194] The mechanism of inhibition appears to be from blocking JAK3 kinase activity at the ATP-binding site.[194]
May mediate immunosuppression via being a selective JAK3 inhibitor
11.4. Virology
A study using the virological strain PR/8/34 (Influenza A) and WS/33 (H1N1) noted that its replication in macrophages (immune cells) was suppressed with 25uM Berberine, which extended to A549 human epithelial cells (potent suppression) but not MDCK cells.[197] The suppression with Berberine had IC50 values of 0.01μM and 0.44μM for these respective viral strains, which outperformed the reference drug amantadine.[197]
12Interactions with Oxidation
12.1. Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a cellular cytoskeleton extending from the nucleus into the cytoplasma; it is sensitive to changes in oxidation and calcium influx and has an 'ER stress response' to maintain homeostasis known as the unfolded protein response.[198]
In cancer cells, Berberine has been at times noted to induce apoptosis secondary to activating an ER stress response (IC50 160mcg/mL in T98G Glioblastoma cells,[199][200] cervical,[201] oral,[202] variable range of 10-100uM in other cancer cell lines[203]).
In other cells, Berberine appears to attenuate inflammation-dependent ER Stress in endothelial cells where apoptotis was 95% reversed at 20uM[204] and was noted to suppress the induction of GRP78/BiP (correlated with ER stress) and less caspase-3 (released from mitochondria after stress to induce apoptosis).[204] HepG2 (liver) cells have also noted an attenuation of ER stress,[205] and a study in mouse macrophages incubated with HIV has also noted a dose-dependent reduction of ER stress.[206]
There appears to be regulatory aspects of Berberine on the the endoplasmic reticulum, inducing oxidant stress in cancer cells while attenuating oxidant stress under some conditions in healthy cells. Exact mechanisms mediating this regulation not yet characterized
12.2. Lipid Peroxidation
In an ex vivo assay of lipid peroxidation, Berberine inhibited lipid peroxidation directly with an IC50 of 72μM, and was found to be active in TNBS-induced colitis in reducing lipid peroxidation in the colon.[189] Lipid peroxidation has also been shown to be reduced in the β-cells of the pancreas following oral ingestion of 150-300mg/kg bodyweight.[207]
Appears to be a direct anti-oxidant, but is quite weak at directly preventing lipid peroxidation; appears to be much more potent in living systems, and possibly not mediated by direct effects
13Interactions with Hormones
13.1. Testosterone
Due to interactions with CYP3A4 (inhibition of which may increase testosterone) and CYP1A2 (Aromatase),[49] it is theoretical that Berberine may increase circulating testosterone levels; this is currently untested in living systems.
Theoretical testosterone boosting properties that are currently not demonstrated
13.2. Estrogen
The incubation of Tamoxifen (1.5uM) and Berberine (16ug/mL) in estrogen responsive MCF-7 breast cancer cells is able to synergistically increase apoptosis.[208] This synergism seems to be related to estrogen receptor antagonists in general, although the mechanism(s) exerted by Berberine is/are currently not known.[208]
13.3. Leptin
One intervention on people newly diagnosed with metabolic syndrome noted that 300mg Berberine taken thrice a day (900mg total) for 12 weeks was able to reduce circulating leptin levels 36% while nonsignificantly raising adiponectin, but the leptin/adiponectin ratio improved from 0.76 to 0.58.[146]
13.4. GLP-1
Glucagon-like peptide 1 (GLP-1) is a peptide hormone known to be secreted from the gut that has blood glucose lowering properties, in part through stimulating insulin secretion[209][210] and may also have a role in proliferating pancreatic β-cells.[211] Berberine has been found to, at the oral dose of 120mg/kg for 5 weeks, increase both GLP-1 and insulin concentrations in streptozotocin-induced diabetic rats (measured postprandially).[212] This study also noted an increase in β-cell population (460% of diabetic control, but still less than half of nondiabetic control) which was attributed to GLP-1.[212]
14Cancer Metabolism
14.1. Mechanisms (General)
Multiple targets exist which may explain anti-cancer effects of berberine. Berberine is known to directly bind to DNA, which is one mechanism by which it can cause cell cycle arrest in multiple human cell cancer lines in vitro,[15] although upregulation of GADD153, a transcription factor involved in apoptosis, may also play a role and has been seen to accompany cell cycle arrest and apoptosis in a human cervical cancer cell line.[201] This mechanism has been observed in one case to occur through downregulating death-domain-associated protein, a key protein which regulates pathways related to cell survival, by binding to its promoter region, which led triggered a cascasde ultimately leading to cancer cell death.[213] Berberine also has been seen to interfere with DNA synthesis in growing ovarian cancer cells by inhibiting two key enzymes in this pathway, dihydrofolate reductase and thymidylate synthase.[214]
Berberine also seems to supress the expression of certain proteins which are anti-apoptotic in cancer cells, such as Mcl-1.[215] Berberine can also affect telomerase activity through multiple mechanisms as well, including the downregulation of the chaperone protein nucleophosmin,[216]through inhibition of human telomerase reverse transcriptase, an essential component of human telomerase,[217] or perhaps even through direct interaction with telomeric DNA.[74]
Another possible mechanism of berberine's efffect on cancer is via JAK3 selective inhibition, as at least one study has noted that berberine could decrease viability in cancer cell lines overexpressing active JAK3 (Ba/F3-JAK3V674A and L540) while not having a significant effect in other cell lines (HDLM-2 and DU145) at the same concentration of 3uM.[194]
Several plausible molecular mechanisms exist which suggest berberine may have anti-cancer effects.
14.2. Proliferation and Angiogenesis
Berberine seems to act as an antiproliferative in several cancer cell lines in vitro through the induction of apoptosis.[15] The main pathways which berberine exerts its antiproliferative effects include the mitochondrial apoptotic pathway, the HER2/PI3K/AKT pathway, the JNK/p38 redox/ROS pathway, and NF-kB, depending on the cell line.[15]
While direct effects on angiogenesis of tumor cells by berberine have not been observed, berberine is known to suppress Mcl-1 in at least one cell line in vitro, which is known to have angiogenic effects.[215]
14.3. Migration and Metastasis
The migration of a human tongue squamous cell carcinoma line has been shown to be reduced by berberine in vitro, and involved the inhibition of several proteins including NF-κB, MMP-2, and MMP-9.[218] Similar effects have been seen in a human lung cancer cell line.[219]
Injection of berberine into a mouse model of metastatic melanoma inhibited tumor nodule formation; the mechanism for this was an associated downregulation of matrix metalloproteases by negatively regulating ERK1/2.[220]
In vitro studies as well as one in vivo mouse model study suggest that berberine may exhibit anti-metastatic effects.
14.4. Autophagy
Autophagy is the process a cell undergoes to degrade cellular components and to produce energy, usually under times of nutrient deficiency.[221] Its relationship to cancer is complex, as a loss of the ability of autophagy could make cells cancerous by knocking out caspase-independent autophagic, or type II, cell death; parodoxically, however, autophagy may also promote tumor survival by giving cancer cells a growth advantage.[221]
Berberine has been shown in vitro to induce autophagic cell death in human liver cancer cells through the distinct increase in Beclin-1, which is one of the main proteins involved in this pathway, as well as an inhibition of mTOR (through MAPK activation and AKT inhibition), which is one of the main regulators of this pathway.[222] Berberine, at least when combined with radiation, has been shown to induce autophagic cell death in lung carcinoma cells, which also led to tumor shrinkage in a xenograft mouse model.[223]
In addition to inducing classical apoptosis, beberine also may induce autophagic cell death in some cancer cell lines.
14.5. Adjunct Therapy Potential
Berberine has been found to enhance the cytotoxicity of Doxorubicin (a chemotherapeutic agent), where cytotoxicity of Doxorubicin was enhanced with a Combination Index[224] of 0.61-0.73, denoting synergism.[225] This study noted that the IC50 values on growth inhibition with Doxorubicin were enhanced from 3.1 and 16.7uM (A549 and HeLa cells) to 1.7 and 1.9uM despite Berberine being relatively weak.[224] Berberine (60mg/kg) has also been found to reduce the hepatotoxicity of Doxorubicin in rats, attenuating the increase in ALT and AST and reducing Doxorubicin-induced liver necrosis by 28%.[226]
14.6. Brain
Berberine has been seen to inhibit the growth of human neuroblastoma cells which express p53 at concentrations of 5μM-100μM in vitro, with cells not expressing this protein being much less sensitive (IC50 > 100μM), suggesting that p53-induced apoptosis in these cells.[227] Berberine also induced apopotosis in glioblastoma cells in vitro with an IC50 of 134μg/mL through the mitochondrial apoptotic pathway.[228]
14.7. Breast
Berberine induces apoptosis in breast cancer cells in vitro at 25μM through the mitochodrial/caspase-dependent pathway.[229]
14.8. Liver
In HepG2 cells and metastatic liver cells MHCC97-L (as well as nasopharyngeal carcinoma cell lines HONE1 and HK1) appears to induce cytotoxicity with IC50 values at 100uM (HepG2) and 250uM (MHCC97-L) via autophagy, as abolishing Atg5 attenuates cytotoxicity.[222] Berberine was noted to dose-dependently increase staining for autolysosomes and autophagosomes, which may be due to induction of Beclin-1 (which binds to and sequesters Bcl-2,[230] a mitochondrial membrane protection protein; reducing Bcl-2 indirectly promotes autophagy[231]) and lead to cancer cell death via mitochondrial capsase release.[222] A dose-dependent inactivation of mTOR was also noted with Berberine, thought to be downstream of Akt.[222]
14.9. Oral
Berberine at 1-100uM concentration can suppress COX-2 induction in stimulated oral cancer cells (OC2 and KB) secondary to inhibiting the ability of AP-1 to induce their induction; Berberine does not directly interact with either COX-1 or COX-2.[190]
14.10. Thyroid
Berberine has been found to inhibit thyroid cancer cells growth in vitro using the cell lines 8505C and TPC1 (anaplastic and papillary, respectively).[232] This study noted dose-dependent reductions in cell proliferation with an IC50 of 10uM, and was involved with inducing apoptosis at G2/M and G0/G1 for 8505C and TPC1 cells, respectively, modulated by increased expression of P27.[232]
14.11. Colon
In two tested colon cancer cell lines (HT-29 and IMCE) as well as a normal cell line (YAMC), Berberine at 50μM is able to suppress over half the cancer cell growth with further inhibition at 100μM and these doses induced apoptosis in cancer cells; suggesting anti-cancer potential of Berberine in colon cancer cells.[233] Apoptosis was only induced in normal colon cells at 200μM, suggesting that cancer cells are sensitized to the effects of Berberine; the cell death is mediated by oxidation (ROS) inducing AIF release from the mitochondria and accumulation in the nucleus and is caspase independent.[233]
14.12. Prostate
Berberine inhibits the growth of p53-expressing prostate cancer cells in vitro at concentrations of 30-100μM, but has a weaker effect against prostate cancer cells lacking p53 (PC-3 cells) (IC50 > 100μM), as the main mechanism by which berberine operates in this cell line seems to be by inducing p53-dependent apoptosis.[234] Another study on PC-3 cells, however, did find that berberine indeed induces apoptosis in these cells through the mitochondrial/caspase-dependent pathway, and is dependent on the generation of reactive oxygen species.[235]
14.13. Immunological Cancers
A human lymphoma cell line (U937) has been seen to undergo apopoptosis when exposed to 50-75μM berberine in vitro[236] through a mitochondrial/caspase-dependent mechanism.[236] Human HL-60 leukemia cells have also been seen to undergo apoptosis induced by berberine exposure above 30μM, alongside an increase in the production of reactive oxygen species.[237]
15Interactions with the Liver (Hepatology)
15.1. Glucose Interactions
In regards to hepatic gluconeogensis (the production of glucose from non-glucose sources; tends to be drastically increased in the state of diabetes which contributes to elevated fasting glucose) Berberine at high doses (380mg/kg daily) for 5 weeks in diabetic rats is able to reduce the activity of the two rate-limiting enzymes in hepatic gluconeogenesis (PEPCK and G6Pase), which was not related to an increase in insulin (normally suppressive of these two genes).[238] This appeared to be related to normalization of FOXO1 activity, which was increased in diabetic rats,[238] and both the protein content and mRNA (genomic transcription) have been noted to be reduced.[36]
Appears to suppress hepatic gluconeogenesis in diabetic rats, a mechanism that would reduce endogenous glucose production and may confer anti-diabetic effects
In human liver cells (in vitro study), 10mcg/mL over 12 hours can cause an increase in insulin receptor content in hepatocytes; this also extends to other tested cell lines (pancreatic, colon, lymphocyte, fibroblast) between a 1.6 to 1.9 fold increase[175] and appears to work in a PKC dependent and dose-dependent manner.[182] Insulin signalling has also been found to be enhanced secondary to reducing endoplasmic reticulum stress,[205] which is secondary to the insulin receptor.
When tested in CEM cells, coincubation of 0.5nM insulin with 10mcg/mL Berberine induces Akt activation to the same degree as 10nM insulin.[175]
Appears to enhance the signalling of insulin in liver cells, both at the receptor level as well as after the receptor.
15.2. Lipids and Cholesterol
The reduction of LDL receptor expression on the liver, reduced by inflammation, can be effectively preserved with 4 weeks ingestion of 30mg/kg Berberine in rats.[122] In vitro 20uM can increase LDLR by 1.8 fold after 4 hours and 3 fold after 24 hours incubation, and concurrently suppressed PCSK9 mRNA by 30% (as well as abolishing the inducing effect of statins on PCSK9);[239] this inhibitory effect of Berberine on statin-induced PCSK9 induction (which downregulates the LDLR) has been noted elsewhere.[240] Berberine appears to suppress PCSK9 at the promoter level secondary to suppressing the activity of its cofactor, HNF1α.[239]
Berberine has been found to reduce the expression of SREBP2 in hepatocytes,[239] an enzyme that induces the activity of genes in cholesterol synthesis.
Berberine can attenuate the increase in SREBP1 and ChREBP activity, and lessened activity of these two proteins resulted in less activity of Fatty Acid Synthase (FAS) mRNA level which may underlie fatty liver therapeutic potential.[238]
Appears to preserve fatty acid and cholesterol metabolism of the liver during states of inflammation and diabetes
One clinical trial in humans with nonalcoholic fatty liver disease who were given either lifestyle interventions alone, or lifestyle interventions plus 15mg pioglitazone daily or 0.5mg berberine thrice daily for 16 weeks found a significant reduction in total cholesterol in the berberine group compared to lifestyle interventions alone and lifestyle changes plus pioglitazone (-0.52mmol/L versus -0.12mmol/L and -0.11mmol/L respectively) primarily due to triglyceride-lowering, with no significant differences in LDL or HDL between the three groups..[167] Another study found improvements in people with the metabolic syndrome, where 0.5g of berberine three times a day for three months resulted in improved triglycerides versus placebo (2.4mmol/L placebo vs. 1.4mmol/L berberine), with no effect on HDL-C.[176]
15.3. Fibrosis
Not too many studies have looked at Berberine as it pertains to liver fibrosis, but 200mg/kg Berberine in rats (32mg/kg human equivalent) appears to have protective effects against CCL4induced toxicity as assessed by weight of the liver, histology, and immunostaining for the fibrotic indicators of α-SMA and TGF-β1.[241] A subsequent in vitro study on stellate cells noted that 10-20ug/mL Berberine inhibited proliferation of these cells by 31.4+/-1.1% to 53.2+/-0.9% associated with an induction of p21 and p27 and inhibition of FOX01 (all concentration-dependent increases) while a subsequent study with 200-400mg/kg Berberine daily for 6 weeks with twice weekly injections of CCL4 also demonstrated hepatoprotective effects, with 400mg/kg trending to outperform 200mg/kg in regards to serum liver enzymes and histological examination.[242]
One comparative study that induced liver fibrosis in rats with CCL4 injections, Bile Duct Ligation, or alcohol induced fibrosis noted that Berberine (120mg/kg) given therapeutically for 7 weeks noted that Berberine exerted protective effects against all three models; compared to 20mg/kg Tauroursodeoxycholic acid (TUDCA from bear bile) and 150mg/kg Silymarin (Milk Thistle bioactive) Berberine was the most potent against CCL4 fibrogenesis (54% reduction of fibrotic area) and Bile Duct Ligation (57%) but least effective against alcohol (35%, where TUDCA and Silymarin reached 73% and 48%).[243]
In regards to Fibrosis specifically, Berberine has fairly potent anti-fibrotic effects comparable to both Milk Thistle and TUDCA, which is fairly remarkable since those two are highly regarded liver health supplements
15.4. Liver Enzymes
Studies that assess liver enzymes note that Berberine can attenuate the increase in liver enzymes seen with streptozotocin-induced diabetes (380mg/kg for 5 weeks)[238] and CCL4 injections (hepatotoxin) by up to 57% (ALT) or 56% (AST) with 200mg/kg Berberine but 50mg/kg also having a significant effect[241][243] and 400mg/kg being the highest reported oral dose in rats which still follows a dose-dependent reduction of liver enzymes.[242]
When looking at the biomarker of liver enzymes (higher levels in serum are indicative of their release from liver cells which were damaged, and thus correlated with liver damage), Berberine appears to confer protection to liver cells in a very general manner but not overly potent, but appears to be highly dose-dependent against CCL4
One clinical trial in humans with nonalcoholic fatty liver disease who were given either lifestyle interventions alone, or lifestyle interventions plus 15mg pioglitazone daily or 0.5g berberine thrice daily for 16 weeks found a significant reduction in AST and ALT in the berberine group compared to lifestyle interventions alone.[167] There was no difference between the berberine and pioglitazone groups.
15.5. Nonalcoholic Fatty Liver Disease
Berberine has been found to mitigate nonalcoholic fatty liver disease (NAFLD) alongside triglyceride exportation for the liver in rats fed a high-fat diet; the mechanism for this was in part reversal of high-fat diet-induced methylation of the microsomal triglyceride transfer protein (MTTP) promotor.[244] MTTP is necessary for creating and exporting ApoB-containing lipoproteins such as VLDL and LDL.[245]
Success in animals led the researchers to examine the effects of berbirine on NAFLD in humans. People with NAFLD were randomized to receive one of three treatments: lifestyle interventions alone, or with 15mg pioglitazone daily or 0.5g berberine thrice daily for 16 weeks.[167] Adding berberine to lifestyle interventions lowered liver fat content by 17.4% versus 11.4% for lifestyle interventions alone, which was a statistically significant difference.[167] The difference between berberine and pioglitazone was of barely insignificant (p = 0.054).[167] The dose of pioglitazone used in the trial was the lowest usual dose for the drug.
One clinical trail in humans with nonalcoholic fatty liver disease showed that adding berberine to lifestyle interventions decreased fat content of the liver significantly, to an extent similar to the drug pioglitazone at its lowest usual dose.
16Interactions with the Intestinal Tract
16.1. Interactions with Glucose Metabolism
One study in rats note that Berberine was associated with increased GLP-1 secretion as measured in both serum and intestines, secondary to increasing mRNA of proglucagon in the intestines; these may add to anti-diabetic effects.[246]
16.2. Diarrhea
Its anti-diarrheal effects may come vicariously through defense against enterotoxins[247] or through lowering the intestinal motility and transit time.[248]
16.3. Ulcerative Colitis
Ulcerative Colitis (UC) is an Inflammatory Bowel Disease but a condition independent of Crohn's Disease that is characterized by chronic, relapsing inflammation of the intestinal tract as well as general dysregulation of the mucosal immune system where this inflammation adversely affects the host (in this case, human diagnosed with IBD).[249][250]
TNBS (2,6,4-trinitrobenzenesulfonic acid) is a research toxin that replicates colitis via disturbing the immune system in the intestines, and Berberine has repeated evidence for protecting from TNBS induced colitis at a dosage range of 15-100mg/kg bodyweight,[251][189][252] with one noting significant benefit at a dose as low as 5mg/kg (for a 150lb human, 55mg).[253] Studies using Dextran Sulfate (DSS), another colitis producing toxin, reproduce these protective effects where 100mg/kg Berberine Chloride for 5 days after DSS where the increases in TNF-α, IFN-γ, KC, and IL-17 (protein content and mRNA) were all markedly reduced to near control levels, associated with less macrophage activity (inflammatory secretion and response to LPS).[188] These rehabilitative effects on colitis have been replicated at doses as low as 10-40mg/kg in another study, where 10 days of supplementation confered marked protection on inflammatory parameters measured.[254]
Repeated rat studies on Colitis, which suggest that Berberine has remarkable protective effects at fairly low oral doses (as the low absorption rate of Berberine requiring higher doses is no longer required as Berberine does not need absorption to reach the colon)
16.4. Colonic Microflora
One study with Berberine has noted changes in microbiota of the colon.[255]
17Interactions with other Organ Systems
17.1. Mouth
One trial in persons with recurrent aphthous stomatitis (canker sores) noted that a gelatin solution containing 5mg/g Berberine applied for 6 days was effective in reducing pain, ulcer size, and exudation over 6 days.[256]
17.2. Pancreas
In regards to the beta cells (β-cells) of the pancreas, which secrete insulin, 150-300mg/kg Berberine given to diabetic rats who were then fed a high fat/carb diet for 16 weeks appeared to normalize insulin and β-cell concentration of the pancreas as well as pancreatic to body weight ratio, to the same degree with the active control of 4mg/kg Rosiglitazone;[207] this study was rehabilitative in nature, giving treatments 16 weeks after disease progression and suggesting β-cell regeneration linked to normalization of oxidative stress (SOD and MDA).[207]
Acutely, incubations of Berberine with 0.5mM Palmitate (known to damage these cells via lipotoxicity) is dose-dependently reduced between 0.1-1uM concentrations of Berberine.[257] Protection against lipotoxicity and preservation of insulin secretion during lipotoxicity has been noted elsewhere,[152]and the reduction in insulin receptor expression that occurs during a high fat diet (rodents) appears to not occur when 200mg/kg Berberine is ingested.[258]
β-cells of the pancrease appear to both be protected from stressors as well as have their regenerative rates enhanced by Berberine, to a potency similar to the anti-diabetic drug Rosiglitazone
Incubation of normal pancreatic β-cells may slightly (nonsignificantly) increase glucose-stimulated insulin secretion (GSIS) in HIT-T15 cells,[257] with statistical significance being reported elsewhere at 1-10mM (HIT-T15 cells)[259] and up to 50mM (MIN6 cells).[260] This effect has failed to be reported in the βTC3 cell line.[261]
One study noted that incubation of Berberine in MIN6 caused somewhat regulatory effects, suppressing the response to high glucose concentrations but preincubation of Berberine for 24 hours causing increased GSIS in response to subsequently administered glucose.[152] It was later noted that Berberine suppressed cAMP-stimulated insulin secretion.[152]
Berberine can activate AMPK in pancreatic cells, which does not appear to suppress GSIS; Berberine appears to suppress GSIS via cAMP.[152] cAMP is elevated by activation of PKA, which is sensitive to calcium influx into β-cells[262][263] or directly by intracellular Ca2+ mobilization.[264] These interactions, seemingly independent of AMPK, may be related to Berberine being an agonist of the GPR40 receptor (induces calcium mobilization upon activation) with an EC50 of 0.76mM.[265]
For the most part, Berberine appears to induce glucose-dependent insulin secretion from pancreatic cells, which may be due to regulating cAMP-induced insulin secretion. This might be related to being an agonist of the GPR40 receptor, a novel anti-diabetic effect
The protein content of the insulin receptor itself may be increased on pancreatic cells at 10mcg/mL in as little as 12 hours in vitro.[175]
17.3. Lungs
Berberine has been implicated in inducing relaxation of the Guinea Pig Trachea with an EC50 of 34.2+/-0.6mcg/mL (in this study, outperformed by canadine and canadaline[266]) and neither xanthine amine congener nor timolol antagonised the effects of Berberine.[266]
17.4. Kidneys (Renal)
In diabetic rats given a high fat diet, oral intake of 50-200mg/kg Berberine daily reduced the kidney:body weight ratio and improved the histological observation at 100-200mg/kg intake (50mg/kg did not seem too effective) with the increase of type IV collagen and TGF-β1 seen in diabetic rats being prevented with all dose of Berberine; protective efficacy of 100-200mg/kg seemed comparable to Enalapril (1mg/kg) and Simvastatin (2mg/kg) used as active controls;[267] this study is replicated in Medline.[268] Protective effects from a high fat diet on renal damage in diabetic mice has been noted elsewhere with 100-200mg/kg Berberine over 8 weeks, with similar protective effects as 250mg/kg Metformin[269] and other studies have noted that reduces fibronectin[270] or matrix accumulation in general[271][272] following oral treatment.
The increase of TGF-β1 (as well as fibronectin) in the kidneys is mediated by high blood glucose, and precedes diabetic nephropathy[273] and its use as a biomarker for disease progression has been suggested.[274] The inhibition of both TGF-β1 and Fibronectin have been observed from glucose-induced induction of these proteins, with 30uM of Berberine being effective nad 90uM being either equally effective or moreso than the reference drug N,N-dimethylsphingosine (known inhibitor of SPK1).[275] Sphingosine Kinase 1 (SPK1) is a rate limiting enzyme that catalyzes Sphingosine to sphingosine 1-phosphate (S1P)[276] and is activated to a high degree in diabetic kidneys.[277]Berberine has been implicated in suppressing the activity of the SPK1 enzyme in vivo in diabetic mice at 300mg/kg daily for 12 weeks[278] and although it has potent inhibitor effects on SPK1 induction from glucose it does not appear to directly interact with the enzyme.[275] and may be secondary to AP-1 inhibition.[275]
Other possible mechanisms of protection to the kidneys that do not appear to be mediated by anti-inflammatory mechanisms (AP-1 and NF-kB inhibition are generally called antiinflammatory) are anti-oxidative and aldose reductase inhibitory potential;[279] this study being replicated in Medline.[280]The induction of the Aldose reductase receptor that occurs during diabetes appears to not occur if treated with 200mg/kg Berberine throughout the diabetic period.[280] Anti-oxidant effects also seem apparent due to normalization of superoxide dismutase (SOD) and MDA (indicator of lipid peroxidation),[280] and in isolated rat kidney homogenates the anti-oxidative effects (against ferrous-ascorbate lipid peroxidation) of Berberine are slightly more than the reference compound butylated hydroxytoluene.[281]
These protective effects may extent to non-diabetic kidneys, as evidence by a study in diet-induced hypertensive rats that recieved renal protection.[282]
Berberine appears to be effective in disconnecting the adverse effects of high blood glucose on the kidneys, and is most well studied in preventing the fibrotic progression of diabetic nephropathy; this appears to be related to anti-inflammatory mechanisms of action
Berberine has been implicated as having antiurolithic effects (reducing kidney stones),[281] due to one of the herbs it is derived from (Berberis Vulgaris) being used traditionally for this purpose.[283]Berberine was found to have slight diuretic effects between the oral doses of 5-20mg/kg (up to 78%) while increasing urinary sodium and potassium but reducing calcium (similar to hydrochlorothiazide).[281]
Oral ingestion of 500mg Berberine thrice a day (1500mg total) for 12 weeks in otherwise healthy humans has failed to note any renal toxicity associated with supplementation.[128]
17.5. Skin
Berberine has interactions with the skin and has potential to be used as a topical agent, as it suppresses lipid synthesis from sebaceus glands[284] and acts like a surfactant in that it increases skin permeability to polar drugs.[285] If used as a skin agent, however, excessive sunlight should be avoided due to berberine's potential to form free radicals when used externally.[286]
17.6. Penis
In isolated corpus cavenosum smooth muscle cells, Berberine at 10-1000uM concentration was able to concentration-dependently reduce oxidative damage induced by H2O2 and preserve both nitric oxide (NO) and SOD content.[287] Lower concentrations of 1-3uM Berberine was able to concentration-dependently increase the expression of eNOS by 40% and 66% with no influence on iNOS[288] and this was shown to increase penile relaxation (a pro-erectile effect) after direct injections of Berberine into rabbit penises.[289] This study noted a dose-dependent (1-5mg/kg) relaxation in phenylephrine precontracted penile strips with 5mg/kg increasing intracavernous pressure (ICP) by 399%.[289] The mechanism of action of Berberine is direct (non-neuronal) and NO dependent as propanolol, atropine, and Phentolamine could not inhibit the contraction while endothelium removal and L-NAME (NO inhibitor) significantly attenuated the responses; it was independent of muscarinic and β-adrenoceptors.[289] Another possible mechanism is reducing the mRNA expression of PDE5 (both A1 and A2 subsets, with more suppression of the latter), the anti-erectile protein that is the molecular target of Viagra,[290] and Berberine has been found to increase both cAMP and cGMP content in isolated penile strips.[291]
This study[289] noted that Paparvine (Papaver somniferum) as active control was more potent acutely, but Berberine appeared to increase penile tumescence for a longer duration of time (reviewed here).[292]
Appears to have pro-erectile effects, which appear to be nitric oxide mediated and long lasting; no studies in living systems that use oral administration (only a direct injection) and practical relevance of the above is unknown
17.7. Testicles
There appears to be protective effects of a synthetic derivative to Berberine in the testicles (CPU86017-RS),[293][294] but currently no studies exist assessing the effects of Berberine itself.
18Other Possible Therapeutic Roles
18.1. Polycystic Ovarian Syndrome
Polycystic Ovarian Syndrome (PCOS) is a condition affecting women that is commonly associated with insulin resistance, where Metformin is a common pharmaceutical to control side-effects. One trial alongside standard therapy (antiandrogens and estrogen) that compared Berberine and Metformin (both at 1500mg in three doses) against placebo over 3 months noted that Berberine was associated with the largest reduction in waist circumference (despite no significant differences in weight) yet no significant differences existed between measures of insulin resistance, blood glucose, or insulin with both being better than placebo.[295]
Limited evidence, but may be as effective as Metformin for reducing biochemical symptoms of PCOS
18.2. Liver Disorders with concurrently High Glucose
Berberine appears to be safe and effective for reducing triglycerides in persons with liver cirrhosis or hepatitis according to preliminary evidence,[296] and a later study using 1g Berberine for 2 months in persons with Hepatitis C (n=18) or Hepatitis B (n=17) with high blood glucose that there were reductions in fasting blood glucose (13.4-17.1%), triglycerides (17.6-19%) and reduced the elevation of liver enzymes (ALT and AST).[175]
19Nutrient-Nutrient Interactions
19.1. Metformin
Metformin is an anti-diabetic pharmaceutical that is known to indirectly activate AMPK, and does so at a similar potency to Berberine (on an oral dosage basis as well as cellular concentration; reviewed in the glucose metabolism section). This has led to an informal designation of Berberine as Herbal Metformin, and at least one meta-analysis on Berberine noted that they were comparable in efficacy for the treatment of Diabetes.[130] It should be noted that not all mechanisms are shared between the two compounds.
Metformin (2.7 fold uptake at 15uM) and Berberine (2.9 fold at 20uM) show additive effects at enhancing glucose uptake into muscle cells (4.1 fold);[297] one study in adipocytes suggested weak synergism.[158]
Metformin and berberine have the potential to interact with each other through interaction with the organic cation transporters 1 and 2; berberine has been shown to inhibit these proteins, which are one of the major ways cells take up metformin in circulation, which could in theory limit the effects of metformin.[298] Indeed, AUC and plasma levels of metformin were higher after an IV bolus of metformin coadministered with berberine in rats.[298]
Of comparable anti-diabetic potential to Metformin (although less studied than Metformin), and may be either additive or synergistic based on some preliminary in vitroevidence. However, cell uptake of metformin may be limited by berberine due to blockade of organic anion transporters, which could in theory limit the efficacy of metformin. More evidence is needed to explore the clinical effects of this interaction.
19.2. Statins
Berberine by itself is implicated in reducing cholesterol levels by up to 25% in persons with high cholesterol, which is though to be through upregulating and preserving the expression of the LDL receptor (which takes up LDL from the blood into tissues).[65] Berberine has been implicated in attenuating increases of PCSK9 (a protein that suppresses expression of the LDL receptor)[122] which may disconnect the side effect of high cholesterol from conditions that induce PCSK9 activity; this includes inflammation[126] (leading to it being referred to as 'the next statin' by some[299]) and statin drugs, where the induction of PCSK9 appears to limit the statin's own ability to reduce cholesterol.[300][301][302] and as such the combination of statin drugs and berberine for the purpose of lipid and cholesterol reduction appears to be synergistic.[303]
Statin drugs reduce circulating lipids and cholesterol, but over time may induce the activity of a protein known as PCSK9 that suppresses the LDL receptor and curbs further benefits; Berberine inhibits this protein induction, and can both exert cholesterol reducing effects itself and may prolong the benefits of statin drugs on cholesterol
19.3. Policosanol and Red Yeast Extract
The combination of Berberine (usually 500mg), Red Yeast Extract (3mg) and Policosanol (10mg) has been used in a few trials in humans together, and has been implicated in weight loss in persons with metabolic syndrome (2kg over 18 weeks),[1] decreases in blood pressure,[1] reductions in insulin resistance,[304] reductions in total cholesterol,[2][3][1] LDL-C,[305] and improved blood flow.[1][304] At least one study noted that 18 weeks of supplementation in persons with metabolic syndrome resulted in less ventricular mass and an improvement in the E/A ratio (from 1.00 ± 0.43 to 1.12 ± 0.30 over 18 weeks), indicative of better heart health.[1]
Despite this combination being used very frequently, the one study to compare the combination against Berberine in isolation suggest that most benefit is secondary to Berberine rather than a possible synergistic relationship.[2]
The combination of the three appears to be used frequently, most likely not synergistic and the observed benefit seems to be mostly related to Berberine itself
It should be noted that Policosanol tends to be marketed as a cholesterol lowering agent with statin-like mechanisms, which tie into the evidence in the former section on PCSK9; however, Policosanol itself may not be a suitable choice for this due to geographical issues with the research that need to be elucidated (can be read up on the Policosanolpage). Red Yeast Extract is a naturally occurring source of Lovastatin[306]
19.4. Sodium Caprate
Sodium Caprate (decanoic acid or capric acid, bound to sodium) a fatty acid found in coconut oil and other sources, has been demonstrated to enhance the absorption of Berberine when both are stimulaneously fed to rats.[36] This enhancement has been repeated elsewhere,[37] where absorption was increased 1.49-3.49 fold depending on where the absorption was measured in the intestines (greater enhancement in the ileum, less in the jujenum). Neither of these studies noted damage to the intestinal mucosa.[36][37]
Pairing Sodium Caprate with Berberine may increase the absorption rates of berberine significantly
19.5. Berberol
Berberol is a mixture of the two herbs Berberis aristata at 588mg (Tree Turmeric, source of Berberine at 85%) and Silybum marianum (Milk Thistle) at 105mg with over 60% flavonolignans.[33] The combination is thought to be synergistic as Berberine has limited absorption due to P-Glycoprotein efflux in the intestines[27] and Silymarin from Milk Thistle inhibits P-Glycoprotein.[307] It should be noted (and can be read up on the Milk Thistle page) that this inhibition is questionably relevant in humans, and the lead author of the Berberol absorption study has patented Berberol.[33]
Berberol was tested in 22 type II diabetics with poor glycemic control using two of the above tablets daily for 90 days noted reductions in HbA1c (-10.6%), Total Cholesterol (-21%), LDL-C (-19.2%), Triglycerides (-44.1%), and an improvement of insulin resistance as assessed by HOMA-R (26.1%).[33]
Brand name for the mixture of Berberine and Milk Thistle, said to increase absorption of Berberine via inhibition of P-Glycoprotein (known to increase absorption of Berberine per se); whether Milk Thistle is the best method is unclear, and the study demonstrated enhanced absorption is against another study on Milk Thistle (where it failed to alter Digoxin kinetics suggesting no inhibition) and also has potential personal investment by the lead author
19.6. Er-Xian
Er-Xian decoction (EXD) is a mixture of several herbs of which three are deemed mandatory; the cortex of Phellodendri Chinensis (Berberine as bioactive), the rhizome of Curculiginis (curculigoside) and Epimedii Folium (source of Icariin, such as Horny Goat Weed).[308] A study in female mice (menopause model) given the bioactives of Berberine (120mg/kg), Icariin (40mg/kg), and Curculigoside (20mg/kg) for 12 weeks noted that the three bioactives were synergistic in preserving bone mass and that they were nonsignificantly less effective than the active control of 1mg/kg nylestriol weekly.[308]
20Safety and Toxicology
20.1. General
One meta-analysis of 14 trials (1068 diabetic patients) given a variable range of 0.5-1.5g berberine for an average period of 12 weeks with or without standard oral hypoglycemic therapy noted that berberine appeared to be associated with more gastrointestinal/abdominal discomfort and mixed effects on stool (with reports of both diarrhea and constipation), most of which were alleviated with dose reduction or dividing the dose into multiple servings a day; no significant differences were noted between placebo/control and Berberine for hypoglycemia in these diabetics.[130]
Despite inhibition of the CYP1A1 enzyme, and reports of plants containing berberine to exert toxic effects, berberine in itself is considered to have low toxic potential.[139][309]
However, one in vitro study has shown that berberine may have serious interactions with macrolide antibiotics such as azithromycin.[310] This study showed that macrolide antibiotics when combined with berberine decrease currents through potassium channels that may lead to long QT syndrome, a possible serious heart toxicity.[310]
A mouse study found that 5 mg/kg injections of berberine could induce skeletal muscle atrophy through increasing atrogin-1 expression (see Skeletal Muscle and Physical Performance section).[61]Whether this effect extends to humans is not currently known.
In most persons, berberine appears to be relatively safe. A potentially serious interaction between macrolide antibiotics such as azithromycin and berberine exists, though.
20.2. Contraindictions
Due (partially) to inhibition of the CYP3A4 enzyme, berberine can adversely interact with Cyclosporin A and increase bioavailability of the latter, which necessitates a lower dosage.[52][51] Berberine can also adversely interact with warfarin, thiopental, and tolbutamide by displacing them from their sites of action and increasing blood toxicity potential.[139] Macrolide antibiotics such as azithromycin and clarithromycin may also interact with berberine and may lead to serious heart problems.[310]
It also displaces bilirubin from albumin at a very high rate, which may be a factor for reported green stools alongside berberin's natural coloration as yellow. Because of the former reactions, however, it should not be used in jaundiced neonates and pregnant women.[139][311]
Berberine can interact with many drugs, and some of these interactions may increase the chances of serious complications. It should also not be used during pregnancy.
Scientific Support & Reference Citations
References
- Effects of a nutraceutical combination on left ventricular remodeling and vasoreactivity in subjects with the metabolic syndrome.
- Cicero AF, Rovati LC, Setnikar I. Eulipidemic effects of berberine administered alone or in combination with other natural cholesterol-lowering agents. A single-blind clinical investigation. Arzneimittelforschung. (2007)
- Marazzi G, et al. Long-term effects of nutraceuticals (berberine, red yeast rice, policosanol) in elderly hypercholesterolemic patients. Adv Ther. (2011)
- Vuddanda PR, Chakraborty S, Singh S. Berberine: a potential phytochemical with multispectrum therapeutic activities. Expert Opin Investig Drugs. (2010)
- Cernáková M, Kostálová D. Antimicrobial activity of berberine--a constituent of Mahonia aquifolium. Folia Microbiol (Praha). (2002)
- Sheng WD, et al. Treatment of chloroquine-resistant malaria using pyrimethamine in combination with berberine, tetracycline or cotrimoxazole. East Afr Med J. (1997)
- Kaneda Y, et al. In vitro effects of berberine sulphate on the growth and structure of Entamoeba histolytica, Giardia lamblia and Trichomonas vaginalis. Ann Trop Med Parasitol. (1991)
- Ghosh AK, Bhattacharyya FK, Ghosh DK. Leishmania donovani: amastigote inhibition and mode of action of berberine. Exp Parasitol. (1985)
- Kaneda Y, Tanaka T, Saw T. Effects of berberine, a plant alkaloid, on the growth of anaerobic protozoa in axenic culture. Tokai J Exp Clin Med. (1990)
- Grycová L, Dostál J, Marek R. Quaternary protoberberine alkaloids. Phytochemistry. (2007)
- Singh IP, Mahajan S. Berberine and its derivatives: a patent review (2009 - 2012). Expert Opin Ther Pat. (2012)
- Krivogorsky B, et al. Structure-activity studies of some berberine analogs as inhibitors of Toxoplasma gondii. Bioorg Med Chem Lett. (2012)
- Avula B, Wang YH, Khan IA. Quantitative determination of alkaloids from roots of Hydrastis canadensis L. and dietary supplements using ultra-performance liquid chromatography with UV detection. J AOAC Int. (2012)
- Chen J, et al. Analysis of major alkaloids in Rhizoma coptidis by capillary electrophoresis-electrospray-time of flight mass spectrometry with different background electrolytes. Electrophoresis. (2008)
- Tillhon M, et al. Berberine: new perspectives for old remedies. Biochem Pharmacol. (2012)
- Wang JR, et al. Formation and conformation of baicalin-berberine and wogonoside-berberine complexes. Chem Pharm Bull (Tokyo). (2012)
- Turner N, et al. Berberine and its more biologically available derivative, dihydroberberine, inhibit mitochondrial respiratory complex I: a mechanism for the action of berberine to activate AMP-activated protein kinase and improve insulin action. Diabetes. (2008)
- Cheng Z, et al. 8,8-Dimethyldihydroberberine with improved bioavailability and oral efficacy on obese and diabetic mouse models. Bioorg Med Chem. (2010)
- Subeki, et al. Antibabesial activity of protoberberine alkaloids and 20-hydroxyecdysone from Arcangelisia flava against Babesia gibsoni in culture. J Vet Med Sci. (2005)
- Schramm A, et al. HERG channel inhibitors in extracts of Coptidis rhizoma. Planta Med. (2011)
- Zhang TT, et al. CPU 86017, p-chlorobenzyltetrahydroberberine chloride, attenuates monocrotaline-induced pulmonary hypertension by suppressing endothelin pathway. Acta Pharmacol Sin. (2005)
- Gao J, et al. Raisanberine protected pulmonary arterial rings and cardiac myocytes of rats against hypoxia injury by suppressing NADPH oxidase and calcium influx. Acta Pharmacol Sin. (2012)
- Pan GY, et al. Inhibitory action of berberine on glucose absorption. Yao Xue Xue Bao. (2003)
- Maeng HJ, et al. P-glycoprotein-mediated transport of berberine across Caco-2 cell monolayers. J Pharm Sci. (2002)
- Chen W, et al. Bioavailability study of berberine and the enhancing effects of TPGS on intestinal absorption in rats. AAPS PharmSciTech. (2011)
- Pan GY, et al. The involvement of P-glycoprotein in berberine absorption. Pharmacol Toxicol. (2002)
- Chae HW, et al. Effect of ion-pair formation with bile salts on the in vitro cellular transport of berberine. Arch Pharm Res. (2008)
- Tsai P, Tsai TH. Simultaneous determination of berberine in rat blood, liver and bile using microdialysis coupled to high-performance liquid chromatography. J Chromatogr A. (2002)
- Qiu W, et al. Effect of berberine on the pharmacokinetics of substrates of CYP3A and P-gp. Phytother Res. (2009)
- Zhang X, et al. Intestinal absorption mechanisms of berberine, palmatine, jateorhizine, and coptisine: involvement of P-glycoprotein. Xenobiotica. (2011)
- Hwang YH, et al. Effects of berberine and hwangryunhaedok-tang on oral bioavailability and pharmacokinetics of ciprofloxacin in rats. Evid Based Complement Alternat Med. (2012)
- Shan YQ, et al. Berberine analogue IMB-Y53 improves glucose-lowering efficacy by averting cellular efflux especially P-glycoprotein efflux. Metabolism. (2012)
- Di Pierro F, et al. Pilot study on the additive effects of berberine and oral type 2 diabetes agents for patients with suboptimal glycemic control. Diabetes Metab Syndr Obes. (2012)
- Zhou Y, et al. Drug-drug interactions between ketoconazole and berberine in rats: pharmacokinetic effects benefit pharmacodynamic synergism. Phytother Res. (2012)
- Reversible Increase in Tight Junction Permeability to Macromolecules in Rat Ileal Mucosa In Vitro by Sodium Caprate, a Constituent of Milk Fat.
- Zhang M, et al. Sodium caprate augments the hypoglycemic effect of berberine via AMPK in inhibiting hepatic gluconeogenesis. Mol Cell Endocrinol. (2012)
- Enhancement of Sodium Caprate on Intestine Absorption and Antidiabetic Action of Berberine.
- Fan D, et al. Enhancement by sodium caprate and sodium deoxycholate of the gastrointestinal absorption of berberine chloride in rats. Drug Dev Ind Pharm. (2012)
- Zhang Y, et al. Treatment of type 2 diabetes and dyslipidemia with the natural plant alkaloid berberine. J Clin Endocrinol Metab. (2008)
- Khin-Maung-U, et al. Clinical trial of berberine in acute watery diarrhoea. Br Med J (Clin Res Ed). (1985)
- Khan AY, Hossain M, Kumar GS. Binding of plant alkaloids berberine and palmatine to serum albumins: a thermodynamic investigation. Mol Biol Rep. (2013)
- Hazra S, Hossain M, Suresh Kumar G. Binding of isoquinoline alkaloids berberine, palmatine and coralyne to hemoglobin: structural and thermodynamic characterization studies. Mol Biosyst. (2013)
- Wang YX, et al. Synthesis and structure-activity relationship of berberine analogues in LDLR up-regulation and AMPK activation. Bioorg Med Chem. (2012)
- Li Y, et al. Bioactivities of berberine metabolites after transformation through CYP450 isoenzymes. J Transl Med. (2011)
- Zuo F, et al. Pharmacokinetics of berberine and its main metabolites in conventional and pseudo germ-free rats determined by liquid chromatography/ion trap mass spectrometry. Drug Metab Dispos. (2006)
- Qiu F, et al. Isolation and identification of urinary metabolites of berberine in rats and humans. Drug Metab Dispos. (2008)
- Pan JF, et al. Identification of three sulfate-conjugated metabolites of berberine chloride in healthy volunteers' urine after oral administration. Acta Pharmacol Sin. (2002)
- Virtanen P, et al. Natural protoberberine alkaloids from Enantia chlorantha, palmatine, columbamine and jatrorrhizine for thioacetamide-traumatized rat liver. Acta Anat (Basel). (1988)
- Zhao Y, et al. The in vitro inhibition of human CYP1A2, CYP2D6 and CYP3A4 by tetrahydropalmatine, neferine and berberine. Phytother Res. (2012)
- Guo Y, et al. Repeated administration of berberine inhibits cytochromes P450 in humans. Eur J Clin Pharmacol. (2012)
- Xin HW, et al. The effects of berberine on the pharmacokinetics of cyclosporin A in healthy volunteers. Methods Find Exp Clin Pharmacol. (2006)
- Wu X, et al. Effects of berberine on the blood concentration of cyclosporin A in renal transplanted recipients: clinical and pharmacokinetic study. Eur J Clin Pharmacol. (2005)
- Budzinski JW, et al. Modulation of human cytochrome P450 3A4 (CYP3A4) and P-glycoprotein (P-gp) in Caco-2 cell monolayers by selected commercial-source milk thistle and goldenseal products. Can J Physiol Pharmacol. (2007)
- Hermann R, von Richter O. Clinical evidence of herbal drugs as perpetrators of pharmacokinetic drug interactions. Planta Med. (2012)
- Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine.
- Chatuphonprasert W, et al. Modulations of cytochrome P450 expression in diabetic mice by berberine. Chem Biol Interact. (2012)
- Cheng Q, et al. Drug-metabolizing enzyme and transporter expression in a mouse model of diabetes and obesity. Mol Pharm. (2008)
- Viollet B, et al. Cellular and molecular mechanisms of metformin: an overview. Clin Sci (Lond). (2012)
- Han Y, et al. Redox regulation of the AMP-activated protein kinase. PLoS One. (2010)
- Cheng Z, et al. Berberine-stimulated glucose uptake in L6 myotubes involves both AMPK and p38 MAPK. Biochim Biophys Acta. (2006)
- Wang H, et al. Atrogin-1 affects muscle protein synthesis and degradation when energy metabolism is impaired by the antidiabetes drug berberine. Diabetes. (2010)
- Liu LZ, et al. The pivotal role of protein kinase C zeta (PKCzeta) in insulin- and AMP-activated protein kinase (AMPK)-mediated glucose uptake in muscle cells.Cell Signal. (2010)
- Kim WS, et al. Berberine improves lipid dysregulation in obesity by controlling central and peripheral AMPK activity. Am J Physiol Endocrinol Metab. (2009)
- Brusq JM, et al. Inhibition of lipid synthesis through activation of AMP kinase: an additional mechanism for the hypolipidemic effects of berberine. J Lipid Res. (2006)
- Kong W, et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat Med. (2004)
- Hypothalamic malonyl-CoA triggers mitochondrial biogenesis and oxidative gene expression in skeletal muscle: Role of PGC-1α.
- Turnley AM, et al. Cellular distribution and developmental expression of AMP-activated protein kinase isoforms in mouse central nervous system. J Neurochem. (1999)
- Neidle S. Human telomeric G-quadruplex: the current status of telomeric G-quadruplexes as therapeutic targets in human cancer. FEBS J. (2010)
- Ou TM, et al. G-quadruplexes: targets in anticancer drug design. ChemMedChem. (2008)
- Monchaud D, Teulade-Fichou MP. A hitchhiker's guide to G-quadruplex ligands. Org Biomol Chem. (2008)
- Arora A, et al. Binding of berberine to human telomeric quadruplex - spectroscopic, calorimetric and molecular modeling studies. FEBS J. (2008)
- Zhang WJ, et al. 9-Substituted berberine derivatives as G-quadruplex stabilizing ligands in telomeric DNA. Bioorg Med Chem. (2007)
- Ma Y, et al. Synthesis and evaluation of 9-O-substituted berberine derivatives containing aza-aromatic terminal group as highly selective telomeric G-quadruplex stabilizing ligands. Bioorg Med Chem Lett. (2009)
- Bazzicalupi C, et al. The crystal structure of human telomeric DNA complexed with berberine: an interesting case of stacked ligand to G-tetrad ratio higher than 1:1. Nucleic Acids Res. (2012)
- Naasani I, et al. FJ5002: a potent telomerase inhibitor identified by exploiting the disease-oriented screening program with COMPARE analysis. Cancer Res. (1999)
- Mahata S, et al. Berberine modulates AP-1 activity to suppress HPV transcription and downstream signaling to induce growth arrest and apoptosis in cervical cancer cells. Mol Cancer. (2011)
- Wang X, et al. The uptake and transport behavior of berberine in Coptidis Rhizoma extract through rat primary cultured cortical neurons. Neurosci Lett. (2005)
- Olmez E, Ilhan M. Evaluation of the alpha-adrenoceptor antagonistic action of berberine in isolated organs. Arzneimittelforschung. (1992)
- Hui KK, et al. Interaction of berberine with human platelet alpha 2 adrenoceptors. Life Sci. (1991)
- Vizi ES, et al. Berbanes: a new class of selective alpha 2-adrenoceptor antagonists. J Med Chem. (1987)
- Zhao X, et al. Screening the bioactive compounds in aqueous extract of Coptidis rhizoma which specifically bind to rabbit lung tissues beta2-adrenoceptor using an affinity chromatographic selection method. J Chromatogr B Analyt Technol Biomed Life Sci. (2010)
- Kulkarni SK, Dhir A. On the mechanism of antidepressant-like action of berberine chloride. Eur J Pharmacol. (2008)
- Berberine produces antidepressant-like effects in the forced swim test and in the tail suspension test in mice.
- Kong LD, Cheng CH, Tan RX. Monoamine oxidase inhibitors from rhizoma of Coptis chinensis. Planta Med. (2001)
- Castillo J, et al. LED fluorescence spectroscopy for direct determination of monoamine oxidase B inactivation. Anal Biochem. (2005)
- Inhibition of Dopamine Biosynthesis by Protoberberine Alkaloids in PC12 Cells.
- Kalalian-Moghaddam H, et al. Hippocampal synaptic plasticity restoration and anti-apoptotic effect underlie berberine improvement of learning and memory in streptozotocin-diabetic rats. Eur J Pharmacol. (2013)
- Reisi P, et al. Treadmill running improves long-term potentiation (LTP) defects in streptozotocin-induced diabetes at dentate gyrus in rats. Pathophysiology. (2010)
- Artola A. Diabetes-, stress- and ageing-related changes in synaptic plasticity in hippocampus and neocortex--the same metaplastic process. Eur J Pharmacol. (2008)
- Ye L, Wang F, Yang RH. Diabetes impairs learning performance and affects the mitochondrial function of hippocampal pyramidal neurons. Brain Res. (2011)
- Bhutada P, et al. Protection of cholinergic and antioxidant system contributes to the effect of berberine ameliorating memory dysfunction in rat model of streptozotocin-induced diabetes. Behav Brain Res. (2011)
- Kulkarni SK, Dhir A. Possible involvement of L-arginine-nitric oxide (NO)-cyclic guanosine monophosphate (cGMP) signaling pathway in the antidepressant activity of berberine chloride. Eur J Pharmacol. (2007)
- Kulkarni SK, Dhir A. sigma-1 receptors in major depression and anxiety. Expert Rev Neurother. (2009)
- Understanding the Molecular Mechanism of Sigma-1 Receptors: Towards A Hypothesis that Sigma-1 Receptors are Intracellular Amplifiers for Signal Transduction.
- Current hypotheses on sigma receptors and their physiological role: possible implications in psychiatry.
- Monnet FP. Sigma-1 receptor as regulator of neuronal intracellular Ca2+: clinical and therapeutic relevance. Biol Cell. (2005)
- Fontanilla D, et al. The hallucinogen N,N-dimethyltryptamine (DMT) is an endogenous sigma-1 receptor regulator. Science. (2009)
- Huang L, et al. Berberine derivatives, with substituted amino groups linked at the 9-position, as inhibitors of acetylcholinesterase/butyrylcholinesterase. Bioorg Med Chem Lett. (2010)
- Jung HA, et al. Anti-Alzheimer and antioxidant activities of Coptidis Rhizoma alkaloids. Biol Pharm Bull. (2009)
- Ingkaninan K, et al. Acetylcholinesterase inhibitors from Stephania venosa tuber. J Pharm Pharmacol. (2006)
- Chen XW, et al. Interaction of herbal compounds with biological targets: a case study with berberine. ScientificWorldJournal. (2012)
- Asai M, et al. Berberine alters the processing of Alzheimer's amyloid precursor protein to decrease Abeta secretion. Biochem Biophys Res Commun. (2007)
- Zhu F, et al. Decrease in the production of β-amyloid by berberine inhibition of the expression of β-secretase in HEK293 cells. BMC Neurosci. (2011)
- Lee B, et al. Phellodendron amurense and Its Major Alkaloid Compound, Berberine Ameliorates Scopolamine-Induced Neuronal Impairment and Memory Dysfunction in Rats. Korean J Physiol Pharmacol. (2012)
- Zhang Q, et al. Hypoxia-inducible factor 1 mediates the anti-apoptosis of berberine in neurons during hypoxia/ischemia. Acta Physiol Hung. (2012)
- Hu J, et al. PI3K p55γ promoter activity enhancement is involved in the anti-apoptotic effect of berberine against cerebral ischemia-reperfusion. Eur J Pharmacol. (2012)
- Kwon IH, et al. Effects of berberine on 6-hydroxydopamine-induced neurotoxicity in PC12 cells and a rat model of Parkinson's disease. Neurosci Lett. (2010)
- Segura Aguilar J, Kostrzewa RM. Neurotoxins and neurotoxic species implicated in neurodegeneration. Neurotox Res. (2004)
- Nagatsu T. Isoquinoline neurotoxins in the brain and Parkinson's disease. Neurosci Res. (1997)
- Lee B, et al. Effect of berberine on depression- and anxiety-like behaviors and activation of the noradrenergic system induced by development of morphine dependence in rats. Korean J Physiol Pharmacol. (2012)
- Chang W, et al. Berberine Attenuates Ischemia-Reperfusion Injury Via Regulation of Adenosine-5'-monophosphate Kinase Activity in Both Non-ischemic and Ischemic Areas of the Rat Heart. Cardiovasc Drugs Ther. (2012)
- Wang LH, et al. Berberine alleviates ischemic arrhythmias via recovering depressed I(to) and I(Ca) currents in diabetic rats. Phytomedicine. (2012)
- Gong LL, et al. Coptisine exert cardioprotective effect through anti-oxidative and inhibition of RhoA/Rho kinase pathway on isoproterenol-induced myocardial infarction in rats. Atherosclerosis. (2012)
- Kim YM, et al. Palmatine from Coptidis rhizoma reduces ischemia-reperfusion-mediated acute myocardial injury in the rat. Food Chem Toxicol. (2009)
- Soloff MS, et al. Cloning, characterization, and expression of the rat relaxin gene. Gene. (2003)
- Gu HP, et al. Up-regulating relaxin expression by G-quadruplex interactive ligand to achieve antifibrotic action. Endocrinology. (2012)
- Du XJ, et al. Cardiovascular effects of relaxin: from basic science to clinical therapy. Nat Rev Cardiol. (2010)
- Salehi S, Filtz TM. Berberine possesses muscarinic agonist-like properties in cultured rodent cardiomyocytes. Pharmacol Res. (2011)
- Effect of berberine on Cdk9 and cyclin T1 expressions in myocardium of diabetic rats.
- Zeng XH, Zeng XJ, Li YY. Efficacy and safety of berberine for congestive heart failure secondary to ischemic or idiopathic dilated cardiomyopathy. Am J Cardiol. (2003)
- Zheng L, et al. Protective effect of berberine on cardiac myocyte injured by ischemia-reperfusion. Sichuan Da Xue Xue Bao Yi Xue Ban. (2003)
- Xiao HB, et al. Berberine inhibits dyslipidemia in C57BL/6 mice with lipopolysaccharide induced inflammation. Pharmacol Rep. (2012)
- Lee S, et al. Berberine-induced LDLR up-regulation involves JNK pathway. Biochem Biophys Res Commun. (2007)
- Extracellular Signal-Regulated Kinase–Dependent Stabilization of Hepatic Low-Density Lipoprotein Receptor mRNA by Herbal Medicine Berberine.
- Duff CJ, Hooper NM. PCSK9: an emerging target for treatment of hypercholesterolemia. Expert Opin Ther Targets. (2011)
- Feingold KR, et al. Inflammation stimulates the expression of PCSK9. Biochem Biophys Res Commun. (2008)
- Inhibition of lipid synthesis through activation of AMP kinase: an additional mechanism for the hypolipidemic effects of berberine.
- Hu Y, et al. Lipid-lowering effect of berberine in human subjects and rats. Phytomedicine. (2012)
- Derosa G, et al. Effects of berberine on lipid profile in subjects with low cardiovascular risk. Expert Opin Biol Ther. (2013)
- Dong H, et al. Berberine in the treatment of type 2 diabetes mellitus: a systemic review and meta-analysis. Evid Based Complement Alternat Med. (2012)
- Fatehi-Hassanabad Z, et al. The antihypertensive and vasodilator effects of aqueous extract from Berberis vulgaris fruit on hypertensive rats. Phytother Res. (2005)
- Chun YT, et al. A biochemical study on the hypotensive effect of berberine in rats. Gen Pharmacol. (1979)
- Lau CW, et al. Cardiovascular actions of berberine. Cardiovasc Drug Rev. (2001)
- Wong KK. Mechanism of the aortic relaxation induced by low concentrations of berberine. Planta Med. (1998)
- Effects of berberine on angiotensin-converting enzyme and NO/cGMP system in vessels.
- Cheng B, et al. Cardiovascular aspects of pharmacology of berberine: I. Alpha-adrenoceptor blocking action of berberine in isolated rat anococcygeus muscle and rabbit aortic strip. J Tongji Med Univ. (1987)
- Bova S, et al. On the mechanism of vasodilating action of berberine: possible role of inositol lipid signaling system. J Pharmacol Exp Ther. (1992)
- Huang CG, et al. Effect of berberine on arachidonic acid metabolism in rabbit platelets and endothelial cells. Thromb Res. (2002)
- Imanshahidi M, Hosseinzadeh H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother Res. (2008)
- Cao JW, et al. Effects of berberine on intracellular free calcium in smooth muscle cells of Guinea pig colon. Digestion. (2001)
- Huang Z, et al. Berberine-induced inhibition of adipocyte enhancer-binding protein 1 attenuates oxidized low-density lipoprotein accumulation and foam cell formation in phorbol 12-myristate 13-acetate-induced macrophages. Eur J Pharmacol. (2012)
- Huang Z, et al. Berberine‑attenuated monocyte adhesion to endothelial cells induced by oxidized low‑density lipoprotein via inhibition of adhesion molecule expression. Mol Med Report. (2013)
- Wang Q, et al. Activation of AMP-activated protein kinase is required for berberine-induced reduction of atherosclerosis in mice: the role of uncoupling protein 2.PLoS One. (2011)
- Effect of berberine on glucose transport in adipocyte and its mechanism.
- Huang C, et al. Berberine inhibits 3T3-L1 adipocyte differentiation through the PPARgamma pathway. Biochem Biophys Res Commun. (2006)
- Yang J, et al. Berberine improves insulin sensitivity by inhibiting fat store and adjusting adipokines profile in human preadipocytes and metabolic syndrome patients. Evid Based Complement Alternat Med. (2012)
- Adipocyte stress: The endoplasmic reticulum and metabolic disease.
- Pham TP, Kwon J, Shin J. Berberine exerts anti-adipogenic activity through up-regulation of C/EBP inhibitors, CHOP and DEC2. Biochem Biophys Res Commun. (2011)
- Bae S, Yoon Y. Anti-Adipogenic Activity of Berberine is Not Mediated by the WNT/β-Catenin Pathway. Phytother Res. (2012)
- Zhou J, Zhou S. Berberine regulates peroxisome proliferator-activated receptors and positive transcription elongation factor b expression in diabetic adipocytes.Eur J Pharmacol. (2010)
- Zhou L, et al. Berberine attenuates cAMP-induced lipolysis via reducing the inhibition of phosphodiesterase in 3T3-L1 adipocytes. Biochim Biophys Acta. (2011)
- Zhou L, et al. Berberine acutely inhibits insulin secretion from beta-cells through 3',5'-cyclic adenosine 5'-monophosphate signaling pathway. Endocrinology. (2008)
- Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr Rev. (2005)
- Basu R, et al. Selective downregulation of the high molecular weight form of adiponectin in hyperinsulinemia and in type 2 diabetes: differential regulation from nondiabetic subjects. Diabetes. (2007)
- Wang Y, et al. Post-translational modifications of adiponectin: mechanisms and functional implications. Biochem J. (2008)
- Li Y, et al. Activation of AMPK by berberine promotes adiponectin multimerization in 3T3-L1 adipocytes. FEBS Lett. (2011)
- Lin H, Li Z. Adiponectin self-regulates its expression and multimerization in adipose tissue: an autocrine/paracrine mechanism. Med Hypotheses. (2012)
- Prabhakar PK, Doble M. Effect of Natural Products on Commercial Oral Antidiabetic Drugs in Enhancing 2-Deoxyglucose Uptake by 3T3-L1 Adipocytes. Ther Adv Endocrinol Metab. (2011)
- Wang SH, et al. Effect of Astragalus polysaccharides and berberine on carbohydrate metabolism and cell differentiation in 3T3-L1 adipocytes. Zhongguo Zhong Xi Yi Jie He Za Zhi. (2004)
- Chen C, Zhang Y, Huang C. Berberine inhibits PTP1B activity and mimics insulin action. Biochem Biophys Res Commun. (2010)
- Cok A, et al. Berberine acutely activates the glucose transport activity of GLUT1. Biochimie. (2011)
- Kim SH, et al. Berberine activates GLUT1-mediated glucose uptake in 3T3-L1 adipocytes. Biol Pharm Bull. (2007)
- Bustanji Y, et al. Berberine potently inhibits protein tyrosine phosphatase 1B: investigation by docking simulation and experimental validation. J Enzyme Inhib Med Chem. (2006)
- Yi P, et al. Berberine reverses free-fatty-acid-induced insulin resistance in 3T3-L1 adipocytes through targeting IKKbeta. World J Gastroenterol. (2008)
- Choi BH, et al. Berberine reduces the expression of adipogenic enzymes and inflammatory molecules of 3T3-L1 adipocyte. Exp Mol Med. (2006)
- Yi P, Lu FE, Chen G. Molecular mechanism of berberine in improving insulin resistance induced by free fatty acid through inhibiting nuclear trascription factor-kappaB p65 in 3T3-L1 adipocytes. Zhongguo Zhong Xi Yi Jie He Za Zhi. (2007)
- Yan HM, et al. Efficacy of Berberine in Patients with Non-Alcoholic Fatty Liver Disease. PLoS One. (2015)
- Pan GY, et al. The antihyperglycaemic activity of berberine arises from a decrease of glucose absorption. Planta Med. (2003)
- Liu SZ, et al. Antihyperglycemic effect of the traditional Chinese scutellaria-coptis herb couple and its main components in streptozotocin-induced diabetic rats. J Ethnopharmacol. (2012)
- Liu L, et al. Berberine suppresses intestinal disaccharidases with beneficial metabolic effects in diabetic states, evidences from in vivo and in vitro study. Naunyn Schmiedebergs Arch Pharmacol. (2010)
- Liu L, et al. Berberine attenuates intestinal disaccharidases in streptozotocin-induced diabetic rats. Pharmazie. (2008)
- Tintu I, et al. An isoquinoline alkaloid, berberine, can inhibit fungal alpha amylase: enzyme kinetic and molecular modeling studies. Chem Biol Drug Des. (2012)
- Ni YX. Therapeutic effect of berberine on 60 patients with type II diabetes mellitus and experimental research. Zhong Xi Yi Jie He Za Zhi. (1988)
- Sterne JA, Gavaghan D, Egger M. Publication and related bias in meta-analysis: power of statistical tests and prevalence in the literature. J Clin Epidemiol. (2000)
- Zhang H, et al. Berberine lowers blood glucose in type 2 diabetes mellitus patients through increasing insulin receptor expression. Metabolism. (2010)
- Pérez-Rubio KG, et al. Effect of berberine administration on metabolic syndrome, insulin sensitivity, and insulin secretion. Metab Syndr Relat Disord. (2013)
- Efficacy of Berberine in Patients with Type 2 Diabetes.
- Berberine-stimulated glucose uptake in L6 myotubes involves both AMPK and p38 MAPK.
- Ma X, et al. Berberine-induced activation of 5'-adenosine monophosphate-activated protein kinase and glucose transport in rat skeletal muscles. Metabolism. (2010)
- Gomes AP, et al. Berberine protects against high fat diet-induced dysfunction in muscle mitochondria by inducing SIRT1-dependent mitochondrial biogenesis.Biochim Biophys Acta. (2012)
- Liu LZ, et al. Berberine modulates insulin signaling transduction in insulin-resistant cells. Mol Cell Endocrinol. (2010)
- Kong WJ, et al. Berberine reduces insulin resistance through protein kinase C-dependent up-regulation of insulin receptor expression. Metabolism. (2009)
- Krawiec BJ, et al. AMP-activated protein kinase agonists increase mRNA content of the muscle-specific ubiquitin ligases MAFbx and MuRF1 in C2C12 cells. Am J Physiol Endocrinol Metab. (2007)
- Wang XH, Jiang SM, Sun QW. Effects of berberine on human rheumatoid arthritis fibroblast-like synoviocytes. Exp Biol Med (Maywood). (2011)
- Evidence for metalloproteinase and metalloproteinase inhibitor imbalance in human osteoarthritic cartilage.
- Hu PF, et al. Protective effects of berberine in an experimental rat osteoarthritis model. Phytother Res. (2011)
- Ivanovska N, Philipov S, Hristova M. Influence of berberine on T-cell mediated immunity. Immunopharmacol Immunotoxicol. (1999)
- Yan F, et al. Berberine promotes recovery of colitis and inhibits inflammatory responses in colonic macrophages and epithelial cells in DSS-treated mice. Am J Physiol Gastrointest Liver Physiol. (2012)
- Lee IA, Hyun YJ, Kim DH. Berberine ameliorates TNBS-induced colitis by inhibiting lipid peroxidation, enterobacterial growth and NF-κB activation. Eur J Pharmacol. (2010)
- Kuo CL, Chi CW, Liu TY. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett. (2004)
- Park JE, et al. Alkaloids from Chelidonium majus and their inhibitory effects on LPS-induced NO production in RAW264.7 cells. Bioorg Med Chem Lett. (2011)
- Huang Z, et al. Suppression of oxLDL-induced MMP-9 and EMMPRIN expression by berberine via inhibition of NF-κB activation in human THP-1 macrophages.Anat Rec (Hoboken). (2012)
- Guan S, et al. Effects of berberine on expression of LOX-1 and SR-BI in human macrophage-derived foam cells induced by ox-LDL. Am J Chin Med. (2010)
- Kim BH, et al. Inhibition of the signalling kinase JAK3 alleviates inflammation in monoarthritic rats. Br J Pharmacol. (2011)
- Papageorgiou AC, Wikman LE. Is JAK3 a new drug target for immunomodulation-based therapies. Trends Pharmacol Sci. (2004)
- Malaviya R, et al. Targeting Janus kinase 3 in mast cells prevents immediate hypersensitivity reactions and anaphylaxis. J Biol Chem. (1999)
- Cecil CE, et al. Inhibition of H1N1 influenza A virus growth and induction of inflammatory mediators by the isoquinoline alkaloid berberine and extracts of goldenseal (Hydrastis canadensis). Int Immunopharmacol. (2011)
- Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem. (2005)
- Chen TC, et al. Involvement of reactive oxygen species and caspase-dependent pathway in berberine-induced cell cycle arrest and apoptosis in C6 rat glioma cells. Int J Oncol. (2009)
- Eom KS, et al. Berberine-induced apoptosis in human glioblastoma T98G cells is mediated by endoplasmic reticulum stress accompanying reactive oxygen species and mitochondrial dysfunction. Biol Pharm Bull. (2010)
- Lin JP, et al. GADD153 mediates berberine-induced apoptosis in human cervical cancer Ca ski cells. Anticancer Res. (2007)
- Lin CC, et al. Berberine induces apoptosis in human HSC-3 oral cancer cells via simultaneous activation of the death receptor-mediated and mitochondrial pathway. Anticancer Res. (2007)
- Sun Y, et al. A systematic review of the anticancer properties of berberine, a natural product from Chinese herbs. Anticancer Drugs. (2009)
- Hao X, et al. Berberine ameliorates pro-inflammatory cytokine-induced endoplasmic reticulum stress in human intestinal epithelial cells in vitro. Inflammation. (2012)
- Wang ZS, et al. Berberine reduces endoplasmic reticulum stress and improves insulin signal transduction in Hep G2 cells. Acta Pharmacol Sin. (2010)
- Zha W, et al. Berberine inhibits HIV protease inhibitor-induced inflammatory response by modulating ER stress signaling pathways in murine macrophages. PLoS One. (2010)
- Zhou J, et al. Protective effect of berberine on beta cells in streptozotocin- and high-carbohydrate/high-fat diet-induced diabetic rats. Eur J Pharmacol. (2009)
- Liu J, et al. Coptis extracts enhance the anticancer effect of estrogen receptor antagonists on human breast cancer cells. Biochem Biophys Res Commun. (2009)
- Doyle ME, Egan JM. Mechanisms of action of glucagon-like peptide 1 in the pancreas. Pharmacol Ther. (2007)
- Meier JJ, Nauck MA. Glucagon-like peptide 1(GLP-1) in biology and pathology. Diabetes Metab Res Rev. (2005)
- Holst JJ, Gromada J. Role of incretin hormones in the regulation of insulin secretion in diabetic and nondiabetic humans. Am J Physiol Endocrinol Metab. (2004)
- Li J1, et al. Berberine represses DAXX gene transcription and induces cancer cell apoptosis. Lab Invest. (2013)
- Marverti G1, et al. Modulation of the expression of folate cycle enzymes and polyamine metabolism by berberine in cisplatin-sensitive and -resistant human ovarian cancer cells. Int J Oncol. (2013)
- Tsang CM, et al. Berberine suppresses tumorigenicity and growth of nasopharyngeal carcinoma cells by inhibiting STAT3 activation induced by tumor associated fibroblasts. BMC Cancer. (2013)
- Wu HL1, et al. Berberine-induced apoptosis of human leukemia HL-60 cells is associated with down-regulation of nucleophosmin/B23 and telomerase activity. Int J Cancer. (1999)
- Fu L1, et al. Berberine Targets AP-2/hTERT, NF-κB/COX-2, HIF-1α/VEGF and Cytochrome-c/Caspase Signaling to Suppress Human Cancer Cell Growth. PLoS One. (2013)
- Ho YT1, et al. Berberine induced apoptosis via promoting the expression of caspase-8, -9 and -3, apoptosis-inducing factor and endonuclease G in SCC-4 human tongue squamous carcinoma cancer cells. Anticancer Res. (2009)
- Peng PL1, et al. Inhibitory effect of berberine on the invasion of human lung cancer cells via decreased productions of urokinase-plasminogen activator and matrix metalloproteinase-2. Toxicol Appl Pharmacol. (2006)
- Hamsa TP1, Kuttan G. Berberine inhibits pulmonary metastasis through down-regulation of MMP in metastatic B16F-10 melanoma cells. Phytother Res. (2012)
- Eisenberg-Lerner A1, Kimchi A. The paradox of autophagy and its implication in cancer etiology and therapy. Apoptosis. (2009)
- Wang N1, et al. Berberine induces autophagic cell death and mitochondrial apoptosis in liver cancer cells: the cellular mechanism. J Cell Biochem. (2010)
- Peng PL1, et al. Synergistic tumor-killing effect of radiation and berberine combined treatment in lung cancer: the contribution of autophagic cell death. Int J Radiat Oncol Biol Phys. (2008)
- Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul. (1984)
- Tong N, et al. Berberine sensitizes mutliple human cancer cells to the anticancer effects of doxorubicin in vitro. Oncol Lett. (2012)
- Zhao X, et al. Protective effects of berberine on doxorubicin-induced hepatotoxicity in mice. Biol Pharm Bull. (2012)
- Choi MS1, et al. Berberine inhibits human neuroblastoma cell growth through induction of p53-dependent apoptosis. Anticancer Res. (2008)
- Eom KS1, et al. Berberine induces G1 arrest and apoptosis in human glioblastoma T98G cells through mitochondrial/caspases pathway. Biol Pharm Bull. (2008)
- Patil JB1, Kim J, Jayaprakasha GK. Berberine induces apoptosis in breast cancer cells (MCF-7) through mitochondrial-dependent pathway. Eur J Pharmacol. (2010)
- Marquez RT, Xu L. Bcl-2:Beclin 1 complex: multiple, mechanisms regulating autophagy/apoptosis toggle switch. Am J Cancer Res. (2012)
- Bcl-2 Inhibition of Autophagy: A New Route to Cancer?.
- Park KS, et al. Berberine inhibited the growth of thyroid cancer cell lines 8505C and TPC1. Yonsei Med J. (2012)
- Wang L, et al. Berberine induces caspase-independent cell death in colon tumor cells through activation of apoptosis-inducing factor. PLoS One. (2012)
- Choi MS1, et al. Berberine inhibits p53-dependent cell growth through induction of apoptosis of prostate cancer cells. Int J Oncol. (2009)
- Meeran SM1, Katiyar S, Katiyar SK. Berberine-induced apoptosis in human prostate cancer cells is initiated by reactive oxygen species generation. Toxicol Appl Pharmacol. (2008)
- Letasiová S1, et al. Berberine-antiproliferative activity in vitro and induction of apoptosis/necrosis of the U937 and B16 cells. Cancer Lett. (2006)
- Lin CC1, et al. Apoptosis of human leukemia HL-60 cells and murine leukemia WEHI-3 cells induced by berberine through the activation of caspase-3.Anticancer Res. (2006)
- Xia X, et al. Berberine improves glucose metabolism in diabetic rats by inhibition of hepatic gluconeogenesis. PLoS One. (2011)
- Li H, et al. Hepatocyte nuclear factor 1alpha plays a critical role in PCSK9 gene transcription and regulation by the natural hypocholesterolemic compound berberine. J Biol Chem. (2009)
- Cameron J, et al. Berberine decreases PCSK9 expression in HepG2 cells. Atherosclerosis. (2008)
- Zhang BJ, et al. Protection by and anti-oxidant mechanism of berberine against rat liver fibrosis induced by multiple hepatotoxic factors. Clin Exp Pharmacol Physiol. (2008)
- Sun X, et al. Berberine inhibits hepatic stellate cell proliferation and prevents experimental liver fibrosis. Biol Pharm Bull. (2009)
- Wang N, et al. A comparative study on the hepatoprotective action of bear bile and coptidis rhizoma aqueous extract on experimental liver fibrosis in rats. BMC Complement Altern Med. (2012)
- Chang X, et al. Berberine reduces methylation of the MTTP promoter and alleviates fatty liver induced by a high-fat diet in rats. J Lipid Res. (2010)
- Wetterau JR, Lin MC, Jamil H. Microsomal triglyceride transfer protein. Biochim Biophys Acta. (1997)
- Lu SS, et al. Berberine promotes glucagon-like peptide-1 (7-36) amide secretion in streptozotocin-induced diabetic rats. J Endocrinol. (2009)
- Sack RB, Froehlich JL. Berberine inhibits intestinal secretory response of Vibrio cholerae and Escherichia coli enterotoxins. Infect Immun. (1982)
- Yuan J, Shen XZ, Zhu XS. Effect of berberine on transit time of human small intestine. Zhongguo Zhong Xi Yi Jie He Za Zhi. (1994)
- Strober W, Fuss I, Mannon P. The fundamental basis of inflammatory bowel disease. J Clin Invest. (2007)
- Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. (2007)
- Zhang M, et al. Evidence for the complementary and synergistic effects of the three-alkaloid combination regimen containing berberine, hypaconitine and skimmianine on the ulcerative colitis rats induced by trinitrobenzene-sulfonic acid. Eur J Pharmacol. (2011)
- Zhou H, Mineshita S. The effect of berberine chloride on experimental colitis in rats in vivo and in vitro. J Pharmacol Exp Ther. (2000)
- Kawashima K, et al. Pharmacological properties of traditional medicine (XXIX): effect of Hange-shashin-to and the combinations of its herbal constituents on rat experimental colitis. Biol Pharm Bull. (2004)
- Hong T, et al. Suppressive effect of berberine on experimental dextran sulfate sodium-induced colitis. Immunopharmacol Immunotoxicol. (2012)
- Zhang X, et al. Structural changes of gut microbiota during berberine-mediated prevention of obesity and insulin resistance in high-fat diet-fed rats. PLoS One. (2012)
- Jiang XW, et al. Effects of berberine gelatin on recurrent aphthous stomatitis: a randomized, placebo-controlled, double-blind trial in a Chinese cohort. Oral Surg Oral Med Oral Pathol Oral Radiol. (2012)
- Gao N, Zhao TY, Li XJ. The protective effect of berberine on β-cell lipoapoptosis. J Endocrinol Invest. (2011)
- Gu JJ, Gao FY, Zhao TY. A preliminary investigation of the mechanisms underlying the effect of berberine in preventing high-fat diet-induced insulin resistance in rats. J Physiol Pharmacol. (2012)
- Leng SH, Lu FE, Xu LJ. Therapeutic effects of berberine in impaired glucose tolerance rats and its influence on insulin secretion. Acta Pharmacol Sin. (2004)
- Ko BS, et al. Insulin sensitizing and insulinotropic action of berberine from Cortidis rhizoma. Biol Pharm Bull. (2005)
- Yin J, et al. Effects of berberine on glucose metabolism in vitro. Metabolism. (2002)
- Leclerc I, Rutter GA. AMP-activated protein kinase: a new beta-cell glucose sensor?: Regulation by amino acids and calcium ions. Diabetes. (2004)
- Activation of protein kinases and inhibition of protein phosphatases play a central role in the regulation of exocytosis in mouse pancreatic beta cells.
- Tsuboi T, et al. Glucagon-like peptide-1 mobilizes intracellular Ca2+ and stimulates mitochondrial ATP synthesis in pancreatic MIN6 beta-cells. Biochem J. (2003)
- Rayasam GV, et al. Identification of berberine as a novel agonist of fatty acid receptor GPR40. Phytother Res. (2010)
- Abdel-Haq H, et al. Relaxant effects of Hydrastis canadensis L. and its major alkaloids on guinea pig isolated trachea. Pharmacol Toxicol. (2000)
- Wang FL, et al. Renoprotective effects of berberine and its possible molecular mechanisms in combination of high-fat diet and low-dose streptozotocin-induced diabetic rats. Mol Biol Rep. (2012)
- Tang LQ, et al. Berberine ameliorates renal injury by regulating G proteins-AC- cAMP signaling in diabetic rats with nephropathy. Mol Biol Rep. (2012)
- Wu D, et al. Ameliorative effect of berberine on renal damage in rats with diabetes induced by high-fat diet and streptozotocin. Phytomedicine. (2012)
- Liu W, et al. Berberine reduces fibronectin and collagen accumulation in rat glomerular mesangial cells cultured under high glucose condition. Mol Cell Biochem. (2009)
- Jiang Q, et al. Berberine attenuates lipopolysaccharide-induced extracelluar matrix accumulation and inflammation in rat mesangial cells: involvement of NF-κB signaling pathway. Mol Cell Endocrinol. (2011)
- Liu W, et al. Effects of berberine on matrix accumulation and NF-kappa B signal pathway in alloxan-induced diabetic mice with renal injury. Eur J Pharmacol. (2010)
- Oh JH, et al. Sequential effects of high glucose on mesangial cell transforming growth factor-beta 1 and fibronectin synthesis. Kidney Int. (1998)
- Mogyorósi A, et al. Utility of serum and urinary transforming growth factor-beta levels as markers of diabetic nephropathy. Nephron. (2000)
- Lan T, et al. Berberine suppresses high glucose-induced TGF-β1 and fibronectin synthesis in mesangial cells through inhibition of sphingosine kinase 1/AP-1 pathway. Eur J Pharmacol. (2012)
- Taha TA, Hannun YA, Obeid LM. Sphingosine kinase: biochemical and cellular regulation and role in disease. J Biochem Mol Biol. (2006)
- Lan T, et al. Sphingosine kinase-1 pathway mediates high glucose-induced fibronectin expression in glomerular mesangial cells. Mol Endocrinol. (2011)
- Lan T, et al. Berberine ameliorates renal injury in diabetic C57BL/6 mice: Involvement of suppression of SphK-S1P signaling pathway. Arch Biochem Biophys. (2010)
- Liu W, et al. Berberine inhibits aldose reductase and oxidative stress in rat mesangial cells cultured under high glucose. Arch Biochem Biophys. (2008)
- Liu WH, et al. Berberine ameliorates renal injury in streptozotocin-induced diabetic rats by suppression of both oxidative stress and aldose reductase. Chin Med J (Engl). (2008)
- Bashir S, Gilani AH. Antiurolithic effect of berberine is mediated through multiple pathways. Eur J Pharmacol. (2011)
- Li HB, et al. Effects and mechanism of berberine on the hypertensive renal injury rats induced by enriched high fat-salt-fructose diet. Zhong Yao Cai. (2011)
- Bashir S, et al. Berberis vulgaris root bark extract prevents hyperoxaluria induced urolithiasis in rats. Phytother Res. (2010)
- Seki T, Morohashi M. Effect of some alkaloids, flavonoids and triterpenoids, contents of Japanese-Chinese traditional herbal medicines, on the lipogenesis of sebaceous glands. Skin Pharmacol. (1993)
- Namba T, et al. Study on baths with crude drug. II.: the effects of coptidis rhizoma extracts as skin permeation enhancer. Yakugaku Zasshi. (1995)
- Inbaraj JJ, et al. Photochemistry and photocytotoxicity of alkaloids from Goldenseal (Hydrastis canadensis L.) 1. Berberine. Chem Res Toxicol. (2001)
- Tan Y, et al. Antioxidant properties of berberine on cultured rabbit corpus cavernosum smooth muscle cells injured by hydrogen peroxide. Acta Pharmacol Sin. (2007)
- Tan Y, et al. Effect of berberine on the mRNA expression of nitric oxide synthase (NOS) in rat corpus cavernosum. J Huazhong Univ Sci Technolog Med Sci. (2005)
- Chiou WF, Chen J, Chen CF. Relaxation of corpus cavernosum and raised intracavernous pressure by berberine in rabbit. Br J Pharmacol. (1998)
- Tan Y, et al. Effect of berberine on the mRNA expression of phosphodiesterase type 5 (PDE5) in rat corpus cavernosum. Zhonghua Nan Ke Xue. (2004)
- Tan Y, et al. Effects of berberine on cyclic GMP and cyclic AMP levels in rabbit corpus cavernosum in vitro. Zhonghua Nan Ke Xue. (2005)
- Drewes SE, George J, Khan F. Recent findings on natural products with erectile-dysfunction activity. Phytochemistry. (2003)
- Liu GL, et al. Endoplasmic reticulum stress mediating downregulated StAR and 3-beta-HSD and low plasma testosterone caused by hypoxia is attenuated by CPU86017-RS and nifedipine. J Biomed Sci. (2012)
- Zhang GL, et al. CPU86017-RS attenuate hypoxia-induced testicular dysfunction in mice by normalizing androgen biosynthesis genes and pro-inflammatory cytokines. Acta Pharmacol Sin. (2012)
- Wei W, et al. A clinical study on the short-term effect of berberine in comparison to metformin on the metabolic characteristics of women with polycystic ovary syndrome. Eur J Endocrinol. (2012)
- Zhao W, et al. Reduction of blood lipid by berberine in hyperlipidemic patients with chronic hepatitis or liver cirrhosis. Biomed Pharmacother. (2008)
- Prabhakar PK, Doble M. Synergistic effect of phytochemicals in combination with hypoglycemic drugs on glucose uptake in myotubes. Phytomedicine. (2009)
- Kwon M, et al. Organic cation transporter-mediated drug-drug interaction potential between berberine and metformin. Arch Pharm Res. (2015)
- Vogel RA. PCSK9 inhibition: the next statin. J Am Coll Cardiol. (2012)
- Rashid S, et al. Decreased plasma cholesterol and hypersensitivity to statins in mice lacking Pcsk9. Proc Natl Acad Sci U S A. (2005)
- Welder G, et al. High-dose atorvastatin causes a rapid sustained increase in human serum PCSK9 and disrupts its correlation with LDL cholesterol. J Lipid Res. (2010)
- Dubuc G, et al. Statins upregulate PCSK9, the gene encoding the proprotein convertase neural apoptosis-regulated convertase-1 implicated in familial hypercholesterolemia. Arterioscler Thromb Vasc Biol. (2004)
- Kong WJ, et al. Combination of simvastatin with berberine improves the lipid-lowering efficacy. Metabolism. (2008)
- Affuso F, et al. Effects of a nutraceutical combination (berberine, red yeast rice and policosanols) on lipid levels and endothelial function randomized, double-blind, placebo-controlled study. Nutr Metab Cardiovasc Dis. (2010)
- Pisciotta L, Bellocchio A, Bertolini S. Nutraceutical pill containing berberine versus ezetimibe on plasma lipid pattern in hypercholesterolemic subjects and its additive effect in patients with familial hypercholesterolemia on stable cholesterol-lowering treatment. Lipids Health Dis. (2012)
- Liu J, et al. Chinese red yeast rice (Monascus purpureus) for primary hyperlipidemia: a meta-analysis of randomized controlled trials. Chin Med. (2006)
- Zhou S, Lim LY, Chowbay B. Herbal modulation of P-glycoprotein. Drug Metab Rev. (2004)
- Xue L, et al. Effects and interaction of icariin, curculigoside, and berberine in er-xian decoction, a traditional chinese medicinal formula, on osteoclastic bone resorption. Evid Based Complement Alternat Med. (2012)
- Vrzal R, et al. Activation of the aryl hydrocarbon receptor by berberine in HepG2 and H4IIE cells: Biphasic effect on CYP1A1. Biochem Pharmacol. (2005)
- Zhi D, et al. The enhancement of cardiac toxicity by concomitant administration of Berberine and macrolides. Eur J Pharm Sci. (2015)
- Chan E. Displacement of bilirubin from albumin by berberine. Biol Neonate. (1993)
Via HEM and FAQ:
- Affuso F, et al. A nutraceutical combination improves insulin sensitivity in patients with metabolic syndrome. World J Cardiol. (2012)
- Bahadoran Z, et al. The Nitrate-Independent Blood Pressure-Lowering Effect of Beetroot Juice: A Systematic Review and Meta-Analysis. Adv Nutr. (2017)
- Lara J, et al. Effects of inorganic nitrate and beetroot supplementation on endothelial function: a systematic review and meta-analysis. Eur J Nutr. (2016)
- Aune D, et al. Fruit and vegetable intake and the risk of cardiovascular disease, total cancer and all-cause mortality-a systematic review and dose-response meta-analysis of prospective studies. Int J Epidemiol. (2017)
- Ried K. Garlic Lowers Blood Pressure in Hypertensive Individuals, Regulates Serum Cholesterol, and Stimulates Immunity: An Updated Meta-analysis and Review. J Nutr. (2016)
- Ried K, Toben C, Fakler P. Effect of garlic on serum lipids: an updated meta-analysis. Nutr Rev. (2013)
- El Asmar MS, Naoum JJ, Arbid EJ. Vitamin k dependent proteins and the role of vitamin k2 in the modulation of vascular calcification: a review. Oman Med J. (2014)
- Knapen MH, et al. Menaquinone-7 supplementation improves arterial stiffness in healthy postmenopausal women. A double-blind randomised clinical trial.Thromb Haemost. (2015)
- . Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. . ()
- Cockayne S, et al. Vitamin K and the prevention of fractures: systematic review and meta-analysis of randomized controlled trials. Arch Intern Med. (2006)
- Kodama S, et al. Fasting and post-challenge glucose as quantitative cardiovascular risk factors: a meta-analysis. J Atheroscler Thromb. (2012)
- Mannucci E, et al. Is glucose control important for prevention of cardiovascular disease in diabetes?. Diabetes Care. (2013)
- Lan J, et al. Meta-analysis of the effect and safety of berberine in the treatment of type 2 diabetes mellitus, hyperlipemia and hypertension. J Ethnopharmacol. (2015)
- Schaffer SW, et al. Physiological roles of taurine in heart and muscle. J Biomed Sci. (2010)
- Ahmadian M, Dabidi Roshan V, Ashourpore E. Taurine Supplementation Improves Functional Capacity, Myocardial Oxygen Consumption, and Electrical Activity in Heart Failure. J Diet Suppl. (2017)
- Ahmadian M, et al. Taurine supplementation has anti-atherogenic and anti-inflammatory effects before and after incremental exercise in heart failure. Ther Adv Cardiovasc Dis. (2017)
- Beyranvand MR, et al. Effect of taurine supplementation on exercise capacity of patients with heart failure. J Cardiol. (2011)
- Sun Q, et al. Taurine Supplementation Lowers Blood Pressure and Improves Vascular Function in Prehypertension: Randomized, Double-Blind, Placebo-Controlled Study. Hypertension. (2016)
- Militante JD, Lombardini JB. Treatment of hypertension with oral taurine: experimental and clinical studies. Amino Acids. (2002)
(Common misspellings for Berberine include berbine, bererine, berberin)
Cite this page
"Berberine," Examine.com, published on 23 March 2014, last updated on 13 July 2018,http://examine.com/supplements/berberine/