Calvin Cycle
The light reactions are only half of photosynthesis
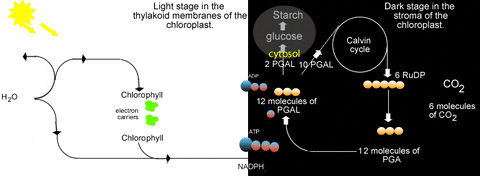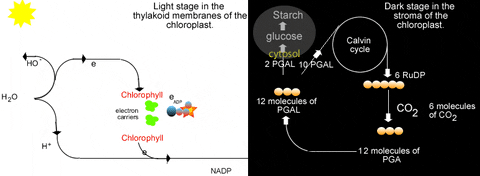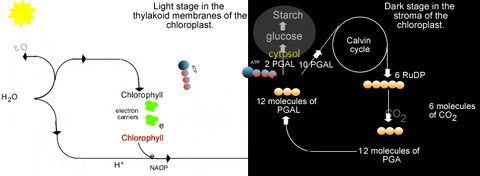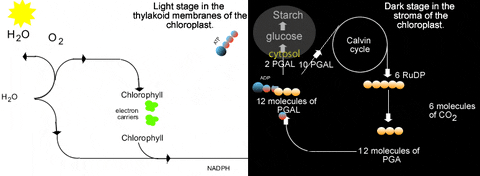
Previously we learned about the light reactions of photosynthesis. In a nutshell these reactions split water to release electrons and produce oxygen gas. The electrons are energized by light energy to higher energy levels in the chlorophyll molecule. They are passed through an electron transfer system in the thylakoid membranes. This is accompanied by proton accumulation in the lumen of the thylakoid which drives ATP synthesis as protons move back to the stroma. The electrons are excited again by light while held in chlorophyll again, and are passed to another short electron transfer system to NADP+. This can be summarized quite briefly as:
| H2O | light → → → → → → → → → chlorophyll | O2 + ATP + NADPH |
|---|
You may recall that photosynthesis can be summarized as:
| CO2 + H2O | light → → → → → → → → → chlorophyll | O2 + CH2O |
|---|
Obviously the light reactions alone do not accomplish photosynthesis. They do not convert carbon dioxide into a carbohydrate which is an integral part of photosynthesis. It is also true that the hydrogen from the water splitting has been attached to a vitamin (NADP+) which is not abundant in cells. There must be a mechanism to use these hydrogens to reduce carbon dioxide into a carbohydrate. This mechanism is generally known as the Calvin cycle after Melvin Calvin in whose laboratory this portion of photosynthesis was dissected. The Calvin cycle has sometimes been referred to as the dark reactions, but these reactions cannot occur in darkness as we shall see. The Calvin cycle has also been called carbon fixation reactions, but only one step of many involves fixing carbon. Although I dislike the "hero worship" that is involved in applying a human name to a botanical reaction, plant physiologists have failed to coin a better term than Calvin cycle for these reactions. Some books use "carbon reactions", but again, not all of the steps involve carbon atoms...and so many other pathways of plant biochemistry involve carbon atoms. So I'm sticking with Calvin cycle.

In order to complement the light reactions and complete photosynthesis, the Calvin cycle obviously can be most-briefly summarized as:
| CO2 + ATP + NADPH | → → → → → → → → → CH2O |
|---|
The Calvin cycle and Light reactions are interdependent
What is obvious from that is the "connection" to the light reactions. The Calvin cycle needs NADPH and ATP from the light reactions. Thus the Calvin cycle is inseparable from the light reactions...they only occur in the light! This is why "dark reactions" is a poor name for the Calvin cycle. This relationship with the light reactions is reciprocal...the light reactions need a supply of ADP, Pi, and NADP+ which come directly from the Calvin cycle:

The Calvin cycle occurs in the stroma
With this knowledge, it is no surprise where the Calvin cycle takes place in a cell. Clearly the need for ATP and NADPH will put the Calvin cycle on the stroma side of the thylakoid membrane where these high-energy molecules are produced by the light reactions. The chemical reactions converting carbon dioxide to carbohydrate are enzymatic rather than electronic, so they are not associated with the thylakoid itself. Rather the Calvin cycle enzymes are dissolved in the stroma (the cytosol of the ancient endosymbiont).
The Calvin cycle is an enzymatic pathway
The enzymes that participate in the Calvin cycle form a cyclic pathway that perpetuates its own raw materials through multiple iterations of the cycle. The pathway is summarized in an overview:

You will notice that carbon dioxide enters the pathway in what is labeled a carboxylation phase. For this phase, the carbon dioxide is added to ribulose-1,5-bisphosphate, a five-carbon sugar-phosphate, bringing the total of carbon atoms to six. The six-carbon intermediate apparently has a very short life and splits to form two 3-phosphoglycerate molecules. 3-PGA is a three-carbon sugar-phosphate. This fact means that the Calvin cycle is sometimes called C-3 photosynthesis too...the product of carbon fixation is a 3-carbon molecule.
Next, the 3-PGA is energized and reduced by ATP and NADPH from the light reactions. This phase is labeled a reduction phase. The reactions here reduce the 3-PGA to make glyceraldehyde-3-phosphate (PGAL aka triose phosphate). This is a three-carbon sugar-phosphate that can be used to make a range of carbohydrates by other pathways (sucrose and starch are just two examples). PGAL siphoned off from this part of the Calvin cycle represents the carbohydrate product of photosynthesis. Yes, I know you probably learned photosynthesis in grade school and were told that C6H12O6 (glucose) was THE product of photosynthesis. That was, of course, a simplification that makes a convenient handle for instruction of children. Now you know that the Calvin cycle product is really the versatile PGAL that can be used dynamically to make a diverse array of carbohydrates. Yup, no Easter Bunny, no Santa Claus, no Tooth Fairy, go Glucose as a photosynthesis product.
Finally the last phase of the Calvin cycle is labeled regeneration as the remaining PGAL is enzymatically converted from the three-carbon sugar-phosphate into more ribulose-1,5-bisphosphate (RuBP) so that the Calvin cycle can continue to go around again. This will of course require an ATP to add the second phosphate, and more than one PGAL to give the total of five carbons found in RuBP.
Each of these three stages of the Calvin cycle (carboxylation, reduction, and regeneration) involve soluble enzymes. The carboxylation step is achieved by the enzyme, ribulose-1,5-bisphosphate carboxylase/oxygenase, also known as RuBisCO (rubisco). This enzyme combines carbon dioxide dissolved in the cytosol with ribulose-1,5-bisphosphate (RuBP) to produce 3-phosphoglycerate (3-PGA):

The carboxylation step is followed by reduction of the 3-PGA to PGAL.

The 3-phosphoglycerate kinase adds a phosphate from ATP to the 3-PGA to make 1,3-bisphosphoglycerate. Then NADP:glyceraldehyde-3-phosphate dehydrogenase removes hydrogen from NADPH and adds it to the 1,3-bisphosphoglycerate to make glyceraldehyde-3-phosphate (PGAL). In this reduction, a phosphate is removed as well. The resulting NADP+, ADP, and Pi can be recycled to the light reactions. These steps demonstrate why the light reactions and Calvin cycle are interdependent. Without the one, the other comes to a complete halt.
Let us remember that at this point we have 6 PGAL, one of which can be used in many ways outside the Calvin cycle to make various sugars, starches, and cellulose for the plant. These may be food for an animal. The sugars can be derivatized in many ways to make other molecules and secondary chemicals for plants. Flavors and fragrances, medicines, lignins, and so on have their origins here. Amino acids and proteins have a basis here too. So while the "traditional" product of photosynthesis is a sugar, we have to keep in mind that a sugar is a versatile material in the biochemical context of a plant cell! More of that below.
If we think about counting carbons, we will need at least three carbon dioxides to enter the cycle to be able to take three carbons off as one PGAL for sugar synthesis, etc. Of course, three cycles of the Calvin cycle would make six PGAL of which one could be used by the cell for various purposes. The other five PGAL (15 carbons total) are needed to regenerate the three RuBP (3⋅5 = 15) that must be regnerated to allow the cycle to run three times. So you need to remember that it takes multiple iterations of the Calvin cycle to allow the production of any useful carbohydrates.

Making PGAL was the easy part of the Calvin cycle. Making RuBP from PGAL is much more interesting. The remainder of the regeneration phase of the Calvin cycle shows how this works.

You will notice that PGAL (glyceraldehyde-3-phosphate) is used in five places to assist in the regneration of RuBP. For two of these, Triose phosphate isomerase converts a PGAL into dihydroxyacetone-3-phosphate. This reaction simply reorganizes the aldehyde into a ketone isomer. The ketone can be combined with various other intermediates to add three-carbons to an expanding sugar molecule.
Aldolase can combine a PGAL and a dihydroxyacetone-3-phosphate to make fructose-1,6-bisphosphate.
Fructose-1,6-bisphosphate phosphatase hydrates the substrate to remove a phosphate and produce fructose-6-phosphate.
We now have a six-carbon sugar-phosphate. Transketolase now combines the fructose-6-phosphate with another PGAL in a reaction involving 9 carbon atoms. These fall apart as a molecule of erythrose-4-phosphate (a four-carbon sugar-phosphate) and a molecule of xylulose-5-phosphate (a five-carbon sugar-phosphate). We will keep the xylulose-5-phosphate in mind for later--it is ready for the last steps in regeneration. But we will focus more on the erythrose-4-phosphate instead for now.
Aldolase combines erythrose-4-phosphate with a dihydroxyacetone-3-phosphate to make sedoheptulose-1,7-bisphosphate. If you are counting carbons correctly, this product has seven carbon atoms and two phosphate groups.
Sedoheptulose-1,7-bisphosphate phosphatase hydrates the seven-carbon sugar-phosphate and removes one of its phosphates. This leaves us with sedoheptulose-7-phosphate...a seven-carbon sugar-phosphate.
Transketolase combines sedoheptulose-7-phosphate with another PGAL to assemble 10 carbons total. These fall apart as a xylulose-5-phosphate and a ribose-5-phosphate...both five-carbons each. We will also recall that we have another xylulose-5-phosphate made earlier.
Ribulose-5-phosphate epimerase will convert xylulose-5-phosphate produced from both previous steps into ribulose-5-phosphate. This enzyme just reorganizes hydroxyl positions in space to convert the 5-carbon sugar-phosphate isomer.
Meanwhile, ribose-5-phosphate isomerase will linearize the ribose-5-phosphate molecule from an earlier step to make an additional ribulose-5-phosphate.
We now have a total of three ribulose-5-phosphate molecules. These are phosphorylated with ATP from the light reactions by ribulose-5-phosphate kinase to make the three ribulose-1,5-bisphosphates needed to regnerate enough Calvin cycle activity to explain the production of one PGAL for the plant cell to use!

The Calvin cycle is a source of complex carbohydrates
Of course, the number of cycles through the Calvin cycle will be much higher than just three. This process is ongoing and iterating repeatedly under good lighting conditions. Because of this, there are small pools of all the intermediates named and diagrammed in the figures on this page!
Because these small pools of intermediates are around at any given moment, any of these molecules may be siphoned off for biochemical use elsewhere. In our regeneration diagram above, you will notice ribose-5-phosphate the sugar-phosphate forming the backbone of RNA. It can be deoxygenated to make deoxyribose-5-phosphate for DNA. You also see sugar-phosphates of 3, 4, 5, 6, and 7 carbons that can form the organic skeletons of a range of secondary molecules. Some of these will be critical in the polymers for cell walls and pectins. Others are for lignin and so on. So anyone who tells you that only glucose is made by photosynthesis is grossly oversimplifying what plants can produce through the Calvin cycle.
We often think of photosynthesis as the source of sucrose and polysaccharides. But technically these products are NOT made by the Calvin cycle. In fact the transport sugar, sucrose, is not even synthesized in the chloroplast. Yet most of the photosynthate is arguably used to make sucrose and starch.
Starch can be synthesized within the chloroplast

The triose phosphates from the Calvin cycle can be combined into fructose-1,6-bisphosphate and fructose-6-phopshate in portions of the regeneration phase.
The fructose-6-phosphate can be converted by the enzyme, hexose phosphate isomerase into glucose-6-phosphate. This, in turn, can be converted to glucose-1-phosphate by the enzyme, phosphoglucomutase. Then ADP glucose pyrophosphorylase can bond AMP to the phosphate of glucose-1-phosphate by removing pyrophosphate from ATP. This produces ADP-glucose, which is the substrate for starch synthase to polymerize into starch.
Pyrophosphate is a very high energy dimer that is converted into two inorganic phosphates by the enzyme pyrophosphatase. The resulting energy drives many reactions in the chloroplast and the cell.
The starch produced inside the chloroplast is obviously a macromolecule. And, because vesicular transport is not known to occur inside chloroplasts, the starch forms around a crystallizing nucleus and forms an amorphous starch grain that respresents a storage polysaccharide. As a chloroplast accumulates much starch, it may become an amyloplast.

Most of the triosephosphate is used to make sucrose for transport
While the chloroplast can become an amyloplast by storing away triose as starch, Most of the chloroplasts in leaves of plants export most of their triose out of the chloroplast and into the cytosol. In the cytosol, the triose is converted into sucrose. The sucrose is then loaded into the phloem for export to the rest of the plant in translocation.

As you can see in the diagram above, in the cytosol, aldolase, fructose-1,6-bisphosphatase, hexose phosphate isomerase, and phosphoglucomutase are present to synthesize fructose-6-phosphate and glucose-1-phosphate. These are the same enzymes found in the chloroplast involved in starch synthesis.
What is different in sucrose synthesis is that UDP-glucose pyrophosphorylase is present and attaches a UMP to the phosphate of glucose-1-phosphate. This produces UDP-glucose which is combined with fructose-6-phosphate by sucrose phosphate synthase. The resulting sucrose phosphate is stripped of its phosphate by sucrose phosphate phosphatase. The product is sucrose that can be loaded into the phloem for transport, and in some cases (sugar cane stems, sugar beet roots) loaded into vacuoles for storage.
Below is a diagram we have seen previously. It shows that the cells of the leaf mesophyll that are responsible for most of photosynthesis have sucrose transport proteins responsible for facilitated diffusion of sucrose out of the cells and into the mesophyll apoplastic fluid. From there, the bundle sheath cells of the leaf vein actively load sucrose into the phloem to generate -ψs, which attracts water from xylem, and increases the pressure potential of the phloem interior. This pressure drives bulk flow from the leaf to the rest of the plant...even bidirectionally from source to sink.

You can also observe above that cells of the root cortex have a proton-sucrose symport protein in the cell membrane to accumulate sucrose from the phloem. This sugar is converted to starch to maintain a sucrose gradient across the root cross section. It is also true that along the pathway from leaf to root, many cells use this kind of antiport to accumulate sucrose in the cytosol. However, in the ground parenchyma cells of sugar cane stems (the cells filling up the interior of the stem around the vascular bundles), to keep this sucrose osmotically separated from cytosol, the vacuole has a proton antiport transporter that allows the cells to accumulate large amounts of sucrose (see below). This tissue we can extract for table sugar. This same kind of situation exists in the roots of sugar beet cortical cells, which are also a source for table sugar for temperate zone farmers.




C is for Chlorophyll
Chlorophyll is Like Liquid Sunshine…
It’s amazing that plants can use chlorophyll to literally turn sunlight into energy via photosynthesis (we’ll spare you the biology lecture). Even molecularly, chlorophyll’s structure is almost identical to hemoglobin’s molecular structure. The only difference is chlorophyll has magnesium as its center while hemoglobin has iron as its center.


Think Green
No wonder why eating our greens is so good for us! Anything that is green is full of chlorophyll (green eggs and ham may be an exception).
Sometimes, we tell customers to try juicing their greens. You can purchase a juicer or just buy your groceries here and we can juice them for you! (It’s just a $1.50 extra) We’ve had customers tell us that celery and carrot juiced together tastes like chocolate milk…guess you’ll have to give that one a shot! Other people love to juice kale, spinach, cucumbers. A little parsley goes a long ways and easy on the fresh beet juice, some people say it’s better to let beet juice sit an hour before drinking it. Of course juicing green colored vegetables are always your safest bet to ensure a megadose of healthy chlorophyll. You can also make green smoothies. Don’t be afraid to experiment for yourself: add some ginger or lemon to add some zest!
Other great sources of chlorophyll are wheatgrass, barley grass, and organic green powders. Martin says the best sources he recommends are Sweet Wheat in capsule form and Evergreen WheatGrass Juice (flash-frozen in cubes on site as soon as they harvest it, found in the freezer section). He also likes BarleyMax by Hallelujah Acres.
No comments:
Post a Comment